Интересное сегодня
Как семантическое соответствие ускоряет мультисенсорное восп...
Введение в мультисенсорное восприятие Мультисенсорные стимулы улучшают восприятие по сравнению с их ...
Что такое размер эффекта и как его интерпретировать в исслед...
Что такое размер эффекта? Размер эффекта — это количественная мера величины экспериментального эффек...
Новое исследование выявляет скрытые паттерны сетевой активно...
Новое исследование, проведенное командой из Университета Джорджии, раскрывает удивительные сведения ...
Готовы ли вы к любви? Понимание готовности к обязательствам ...
Введение Вы действительно готовы к любви? Многие задумываются о том, как найти долгосрочного партнер...
Как цифровые технологии влияют на двойные инновации компаний...
Введение С развитием цифровой экономики такие технологии, как искусственный интеллект, облачные вычи...
Почему люди стремятся к власти: психология альтруизма и эгои...
Почему одни стремятся к власти, а другие помогают людям? Для тех, кто не заботится о власти, наблюде...

Состояние мозга в покое и моторные навыки: исследование быстрой динамики
Моторное обучение (МО) — это динамичный процесс, который обеспечивает эффективное выполнение повседневных действий. Обычно МО требует множества повторений для достижения быстроты и точности движений. Быстрые изменения в производительности наблюдаются во время практики (улучшение «на лету»), а более медленные улучшения происходят вне практики (офлайн) в периоды после тренировки¹. Затем следует перестройка связанных с этим нейронных сетей.
Гипотеза исследования: мы предположили, что быстрые, кратковременные (менее секунды) колебания активности нейронных сетей, регистрируемые с помощью магнитоэнцефалографии (МЭГ) в состоянии покоя (СП), отражают ранние механизмы пластичности, связанные с обучением, и/или меж индивидуальные различия в моторной производительности.
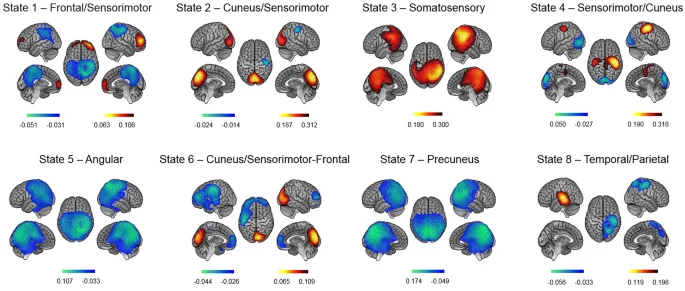
Была записана активность мозга в состоянии покоя (МЭГ) до и через 20 минут после моторного обучения. Скрытое марковское моделирование (СММ) сигналов огибающей мощности МЭГ выявило 8 повторяющихся топографических состояний. Для двух состояний уровни моторной производительности были связаны с временными параметрами СММ как в сессиях состояния покоя до, так и после обучения. Однако, ассоциаций с офлайн изменениями производительности не обнаружено. Эти результаты предполагают устойчивую связь между спонтанной кратковременной нейронной динамикой в состоянии покоя и меж индивидуальными различиями в моторных способностях. С другой стороны, кратковременная динамика состояния покоя, по-видимому, не зависит от состояния, то есть не модулируется опытом обучения, и отражает нейронную пластичность, по крайней мере, на короткой временной шкале.
Введение в моторное обучение
Моторное обучение (МО) — это динамичный процесс, который обеспечивает эффективное функционирование в повседневной жизни. Как правило, МО требует множества повторений для облегчения быстрого и точного выполнения движений. Во время практики моторных задач наблюдаются быстрые изменения производительности (улучшение «на лету»), в то время как более медленные улучшения происходят вне практики (офлайн) в периоды после тренировки¹‚². Динамические изменения производительности во времени предполагают, что МО проходит через критические периоды в рамках этой быстрой/медленной структуры. Действительно, офлайн эволюция моторной производительности характеризуется спонтанным улучшением через 30 минут после практики (период «буста»), которое больше не наблюдается при тестировании через несколько часов (период «молчания» )³,⁴, после чего производительность остается на уровне конца обучения. Дальнейшее увеличение производительности происходит в течение ночи. Было обнаружено, что уровни производительности, достигнутые на ранней, кратковременной фазе «буста», предсказывают офлайн улучшение производительности через 48 часов⁴, что указывает на функциональную значимость непосредственных периодов после тренировки для быстрой реорганизации нейронных сетей, поддерживающих МО и его долгосрочную консолидацию¹‚²‚³‚⁴.
Нейровизуализация и моторное обучение
Исследования нейровизуализации дали представление о нейроанатомических основах МО и их функциональных взаимодействиях в соответствующих сетях мозга. В настоящее время меры функциональной связности позволяют исследовать архитектуру функциональных сетей мозга как во время фактической практики МО (функциональная связность на основе задач), так и в «неактивные» периоды до и после обучения (функциональная связность в состоянии покоя [СП]) . Особенности сетей мозга во время практики задач, такие как гибкость⁵, длина локальных путей, сила связности и эффективность узлов⁶, не только изменяются в результате МО, но и предсказывают будущие уровни обучения⁵. Кроме того, функциональная связность сетей, полученная из измерений СП до моторной задачи, предсказывает индивидуальные способности к МО⁷, в то время как характеристики СП после обучения были предложены как отражающие индуцированную задачей пластичность⁶‚⁸. Многие исследования подробно изучали пространственные компоненты мозга и их взаимодействие в МО, но его основные временные нейронные динамики до сих пор плохо изучены.
Временная динамика нейронной активности
Временная динамика преимущественно исследуется с использованием электрофизиологических методов, таких как магнитоэнцефалография (МЭГ) и электроэнцефалография (ЭЭГ), которые позволяют непосредственно измерять нейронную активность с высоким временным разрешением до миллисекунд, в отличие от функциональной магнитно-резонансной томографии (фМРТ), которая косвенно измеряет нейронную активность через BOLD (blood oxygenation level dependent) отклик мозга в масштабе секунд. Быстрые колебания нейронной активности, улавливаемые с помощью электрофизиологических мер⁹‚¹⁰‚¹¹‚¹², выявили богатую пространственно-временную динамику¹³‚¹⁴‚¹⁵ в сетях состояния покоя (RSN). Например, Бейкер и коллеги¹⁶, используя скрытое марковское моделирование (СММ) огибающих сигналов мощности восстановленных данных МЭГ, показали, что паттерны активности в RSN изменяются гораздо быстрее, чем считалось ранее. Действительно, этот анализ СММ, а также более поздние исследования¹⁷‚¹⁸‚¹⁹‚²⁰‚²¹‚²², выявили дискретные переходные (100–200 мс) состояния мозга, которые повторяются во времени в нейронной активности, связанной с СП, и соответствуют активации/деактивации известных RSN¹⁴‚²³‚²⁴. В совокупности исследования СММ поддерживают гипотезу о том, что нейрокогнитивные сети адаптируются к быстро меняющимся вычислительным требованиям когнитивной обработки²⁵ через механизмы быстрой реорганизации и координации, действующие в масштабе времени менее одной секунды²⁶. Следовательно, способность СММ использовать превосходное временное разрешение сигналов ЭЭГ/МЭГ открывает интересные перспективы для исследования динамики нейронной пластичности, лежащей в основе МО и его консолидации.
Существующие исследования и цели данного исследования
Насколько нам известно, одно исследование МЭГ с использованием СММ сообщило о специфических изменениях активности бета-диапазона, связанной с движением, во время фактической практики самопроизвольной последовательной визуально-моторной задачи²⁷. Другие исследования МЭГ, специально посвященные функциональной связности сетей мозга в течение длительного времени (т. е. 5 минут), обнаружили, что сенсомоторная связность предсказывает уровни обучения в последующей задаче МО²⁸‚²⁹ и офлайн изменения модуляции мубета в сенсомоторной коре после МО³⁰. Это указывает на то, что меж и внутрииндивидуальные различия в офлайн минутной функциональной связности сетей могут отражать способности к обучению и механизмы нейронной пластичности. Тем не менее, эти исследования не дают информации о быстрой (100–300 мс) динамике активации RSN, которая сопровождает пластичность, связанную с МО, и его консолидацию. В этом контексте настоящее исследование МЭГ с использованием СММ было направлено на проверку того, отражают ли внутрииндивидуальные быстрые изменения нейронной динамики от пре- до пост-обучающей сессии СП состояния, связанные с консолидацией МО, механизмы нейронной пластичности. Во-вторых, мы исследовали, предсказывает ли быстрая переходная динамика сетей в спонтанной активности человеческого мозга в пре-обучающем СП индивидуальные уровни производительности, достигнутые в последующей сессии обучения, и, таким образом, может ли она рассматриваться как устойчивый маркер индивидуальной способности к МО.
Результаты исследования
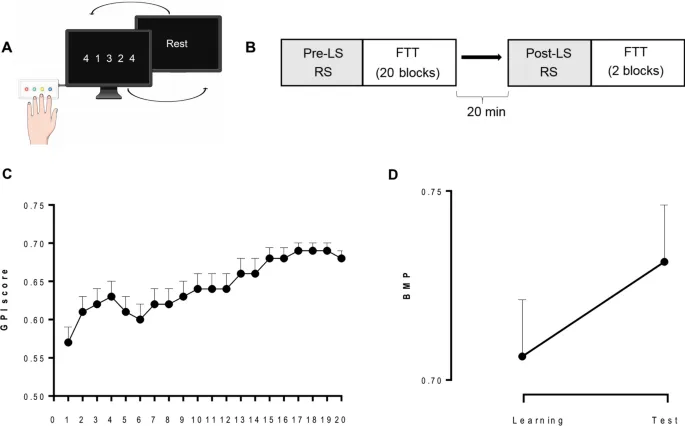
Двадцать семь молодых здоровых участников обучались задаче на моторное запоминание последовательности постукиваний пальцами (FTT1,4) (Рис. 1A,B). Активность мозга записывалась во время пре- и пост-обучающих сессий СП (5 минут, глаза открыты), а также во время обучения (данные не сообщаются) с использованием 306-канального МЭГ-магнитометра Triux. Пост-обучающая запись СП сопровождалась вторым тестом FTT, проведенным через 20 минут после окончания МО, т. е. в период «буста».
Сонливость и утомляемость
До выполнения моторной задачи оценивались уровни сонливости и утомляемости участников с использованием визуальных аналоговых шкал. Значимых различий между сессиями обучения и тестирования по сонливости (t (26) = −1,16, p = 0,26), а также по показателям утомляемости (t (26) = 1,58, p = 0,13, парный t-тест, дополнительная информация, Таблица 1) не обнаружено.
Производительность моторного обучения
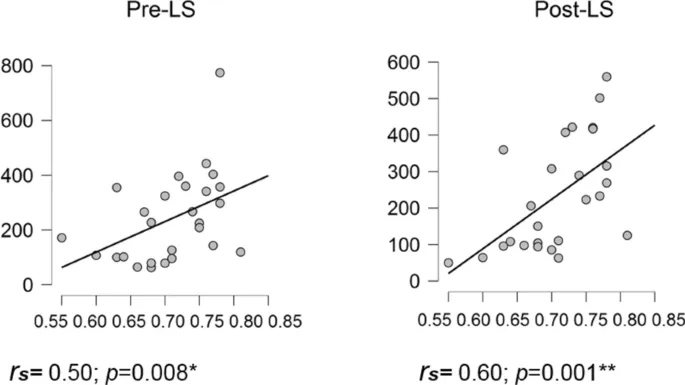
Производительность FTT оценивалась на основе Индекса Глобальной Производительности (GPI), который учитывает компромисс между скоростью и точностью при воспроизведении моторной последовательности³¹. (Для анализа скорости и точности отдельно см. Дополнительную информацию). Во время сессии обучения (LS) производительность быстро улучшалась, достигая асимптотических уровней к концу практики (Рис. 1C). Для каждого индивида лучшая моторная производительность (BMP) при обучении вычислялась путем усреднения двух блоков с наивысшими показателями GPI. Индекс Обучения (LI; улучшение производительности от 2 первых блоков обучения до уровня BMP) был значительным (LI = 23,1 ± 3,3%; одновыборочный t-тест t(26) = 7,0; p < 0,001), что указывает на то, что вышеупомянутые ассоциации для состояний 1 и 6 не зависят от механизмов реорганизации, связанных с офлайн обучением. Что касается состояния 2, то не было выявлено никакой ассоциации между BMP и какими-либо временными параметрами (см. Дополнительную информацию, Рис. 4; все rs < 0,54).
Дополнительно мы провели тот же анализ для всех состояний СММ, независимо от значимых изменений от пре- до пост-обучающей сессии. Наши результаты дополнительно выявили корреляцию между BMP и MLT состояния 7 в пост-обучающей сессии (rs = 0,62, p < 0,02 без коррекции), и коэффициенты корреляции не были значительно отличны между пре- и пост-обучающей сессиями. Аналогично, ассоциации между LI и изменениями временных характеристик состояния от пре- до пост-практики были незначительными (все p > 0,03 без коррекции, Дополнительная информация, Таблица 4). Наконец, мы искали ассоциации между офлайн изменениями производительности (т. е. BE; от лучшей производительности во время обучения до тестовой сессии через 20 минут, см. Дополнительную информацию, Таблица 5). Среднее время жизни состояния 3 (MLT) в пост-обучающей сессии положительно коррелировало с эффектом «буста» (BE), rs = 0,48, p < 0,012 (Дополнительная информация, Рис. 6). Однако после применения более строгой коррекции на множественные сравнения с коэффициентом 28 [число временных параметров и число состояний (4 × 7)], эта корреляция не выдержала статистического порога.
Наконец, мы исследовали, отражено ли моторное обучение в изменениях паттерна переходов между состояниями, т. е. мы вычислили матрицу вероятностей переходов¹⁶ и сравнили переходы до и после обучения (Дополнительная информация, Рис. 8). Однако ни один результат не прошел статистический порог (скорректированный для 8 состояний × 7 вероятностей).
Обсуждение
Данное исследование с использованием СММ сигналов огибающей мощности МЭГ исследовало, предсказывает ли быстрая динамика переходных сетей в спонтанной активности человеческого мозга индивидуальные моторные способности, и отражают ли пост-обучающие изменения быстрой нейронной динамики механизмы пластичности мозга, связанные с обучением. Результаты показывают ассоциацию между уровнями моторной производительности, достигнутыми во время обучения, и временной стабильностью Фронтально/Сенсомоторного Состояния 1 и Состояния 6 (клиновидная/сенсомоторная/фронтальная), как во время пре-, так и пост-моторных сессий СП, предполагая устойчивую связь между спонтанной организацией быстрых рекуррентных сетей мозга, участвующих в моторном обучении, и моторными способностями. Отсутствие корреляций с поведенческими показателями пластичности мозга, связанной с обучением, предполагает, что такая переходная динамика может не отражать изменения нейронных состояний, лежащих в основе моторного обучения и консолидации, по крайней мере, в коротком временном масштабе ранних механизмов консолидации.
Пластичность мозга и временные масштабы
Мы предположили, что пост-обучающие изменения в нейронной динамике будут отражать механизмы консолидации памяти, связанные с офлайн обучением мозга, инициированные в коротком временном окне после окончания обучения²‚⁴. Хотя поведенческие данные, как и ожидалось, продемонстрировали улучшение производительности во время обучения (LI) и офлайн «буст» (BE) через 20 минут после окончания обучения, ни один из этих поведенческих показателей пластичности мозга, связанной с обучением, не был связан с какой-либо динамикой состояний. Такое отсутствие ассоциации может быть связано с тем, что пластические изменения, поддерживающие моторное обучение, одновременно задействуют локальные и удаленные области мозга в иерархии временных масштабов от субсекунд до часов. Исследования фМРТ⁸ и МЭГ²⁸‚³⁰ осветили нейронные сети, связанные с моторным обучением и консолидацией памяти, используя меры функциональной связности в состоянии покоя (rsFC). Основное предположение, лежащее в основе этих методов, заключается в том, что временная корреляция спонтанных сигналов мозга между пространственно распределенными областями мозга создает временно стабильные (по крайней мере, в течение нескольких секунд) функциональные сети. СММ исследует быструю динамику активации этих сетей, и особенно их временную стабильность, в гораздо более коротких временных масштабах, что может препятствовать захвату устойчивых пластических изменений, происходящих в более медленных временных масштабах. Было высказано предположение, что доля времени, проводимого субъектом в каждой сети мозга и мета-состояниях (иерархически организованных состояниях мозга), является последовательной индивидуальной мерой³⁶, генетически обусловленной и демонстрирующей значимые связи с когнитивными чертами. Таким образом, нейропластические изменения могут происходить в пределах определенных границ адаптации в каждой конкретной области мозга. Кроме того, нейропластичность может стимулировать развитие нейронных связей, которые облегчают скорость и точность выполнения, без значительного влияния на временную стабильность сети в целом³⁷, тем самым делая изменения консолидации скрытыми от СММ состояний активности МЭГ.
Индивидуальные особенности или пластические изменения?
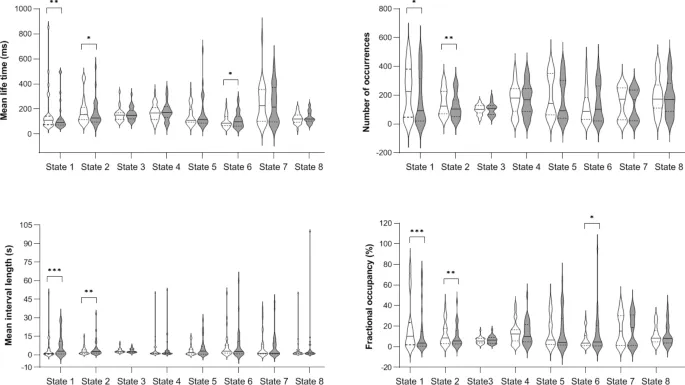
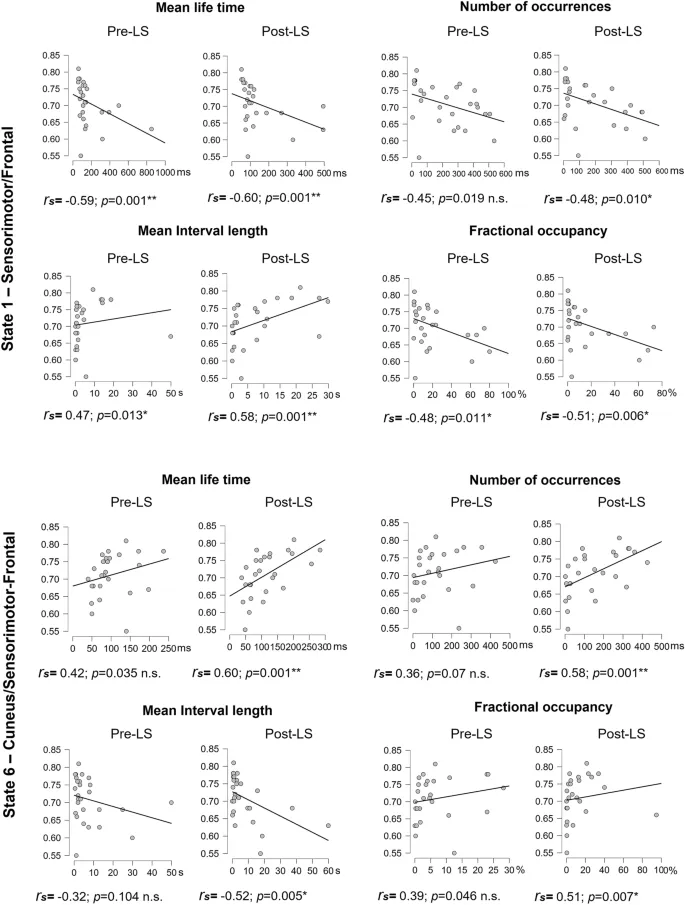
Полученные результаты ставят вопрос о том, являются ли наблюдаемые конфигурации СММ врожденными, стабильными паттернами нейронной активности для данного индивида, т. е. нейронной чертой, или могут ли они модулироваться опытом и требованиями окружающей среды, т. е. нейронными состояниями, отражающими механизмы нейронной пластичности, потенциально связанными с офлайн обучением и процессами консолидации. Примечательно, что предыдущие исследования СММ в основном использовали межгрупповые дизайны для выявления различий между популяциями²⁰‚³⁸‚³⁹. Насколько нам известно, настоящее исследование является первым, которое исследует эволюцию параметров СММ в внутрисубъектном дизайне во время пре- и пост-моторных сессий СП, учитывая уровни моторной производительности и изменения, связанные с обучением. В нашем исследовании СММ выделила 8 переходных рекуррентных состояний из данных огибающих мощности МЭГ в состоянии покоя, имеющих довольно схожие пространственные и временные параметры с предыдущими отчетами¹⁷‚²⁰‚⁴⁰. Количественные временные параметры, измеряющие стабильность и рекуррентность состояний СММ, изменились от пре- до пост-обучающих сессий в Состояниях 1, 2 и 6, что может быть связано с задачей между двумя сессиями СП, или альтернативно, с временем, прошедшим между двумя приобретениями СП. Учитывая первое объяснение, дестабилизация Фронтально/Сенсомоторного Состояния 1 после моторного обучения может быть объяснена автоматизацией практикуемого моторного акта, в конечном итоге требующей одновременно более низкого уровня когнитивного контроля (т. е. меньшей активации фронтальной контрольно-исполнительной сети) и более высокого вовлечения моторных областей (т. е. меньшей деактивации сенсомоторной сети), способствующей консолидации моторной памяти. Такой аргумент нашел бы поддержку в предыдущих исследованиях, показывающих, что обучение моторным навыкам на ранней стадии зависит от активности префронтальных коры, которые, в свою очередь, информируют нижние моторные уровни (т. е. премоторную и первичную моторную кору⁴¹). С прогрессом обучения кортикальная активность во фронтальной доле уменьшается вдоль оси передне-задней⁴²‚⁴³‚⁴⁴‚⁴⁵‚⁴⁶, что позволяет перейти от исполнительного к моторному уровню контроля и, как следствие, снизить требования к центральным ресурсам. Однако наши корреляционные анализы показали, что лучшая моторная производительность (BMP), достигнутая во время сессии обучения, сильно коррелировала с временными параметрами Состояния 1 как в пре-, так и в пост-СП сессиях, без различий между двумя сессиями. Это скорее предполагает существование стабильного, устойчивого паттерна нейронной активности, связанного с индивидуальной способностью к моторной производительности, вывод, дополнительно подкрепленный тем фактом, что мы обнаружили аналогичные паттерны корреляции при проведении корреляций с производительностью в двух первых блоках (базовый уровень, BL) сессии обучения, в течение которой последовательность для изучения еще не интегрирована на процедурном уровне. Другими словами, можно предположить, что повышенная мощность во фронтальных исполнительных сетях в сочетании со сниженной мощностью в сенсомоторных областях может быть важной предпосылкой для успешного выполнения моторной задачи и/или указывать на хорошие индивидуальные моторные способности. С другой стороны, это может быть результатом предыдущего опыта и длительных тренировок в течение развития индивида в моторных задачах, таких как спорт, видеоигры, хобби, связанные с точной моторикой, и т. д.
Различия между состояниями и их интерпретация
С другой стороны, не было обнаружено корреляции между Состоянием 2 и BMP, хотя параметры СММ различались между сессиями, предполагая, что изменения динамики Состояния 2 от пре- до пост-обучения были независимы от моторных эффектов. В отличие от этого, наблюдалась тенденция к положительной ассоциации между BMP и параметрами Состояния 6 в пре-обучающей сессии СП. В обоих состояниях 2 и 6 мощность в клиновидных областях увеличивалась, а в сенсомоторных кортах уменьшалась при активации состояния. Функциональные связи между клиновидной и соматосенсорной областями ранее предполагались⁴⁷, с предположением, что корреляция между этими областями мозга в состоянии покоя способствует антикоррелированной активности, сообщаемой между сетью пассивного режима (метаболически активированной в состоянии покоя⁴⁸) и другими сетями мозга, активированными в контексте задач, требующих визуальной/сенсорной обработки. Хотя одно исследование выявило усиление функциональной связности в пределах сенсомоторных и визуальных RSN вскоре после обучения последовательности⁸, корреляций с производительностью не сообщалось.
Состояние 6 также выявило паттерн деактивации во фронтальной исполнительной сети, противоположный состоянию 1, что может быть ключом к объяснению направленности ассоциаций между уровнями моторной производительности и их временными параметрами. Действительно, лучшие уровни производительности были отрицательно коррелированы с временными параметрами в Состоянии 1 и положительно в Состоянии 6, в соответствии с потенциальным высвобождением исполнительной сети после моторного обучения. Тенденция к стабилизации Состояния 6 после обучения, и, следовательно, тот факт, что его моторные области проводят больше времени будучи деактивированными, также согласуется со сниженной активностью в M1 на поздних стадиях обучения⁴⁹ и автоматизации⁴⁵. Хотя измененная мощность в пределах сенсомоторных кортикальных зон в состояниях может рассматриваться как показатель пластических изменений, связанных с обучением, наши результаты корреляции не подтверждают эту интерпретацию, поскольку ассоциации между паттернами мозговой активности и моторной производительностью схожи до и после эпизода обучения. Следовательно, наши текущие результаты в пользу гипотезы о том, что быстрая динамика мозга отражает, возможно, внутреннюю (т. е. независимую от задачи), устойчивую архитектуру мозга, связанную со способностью выполнять моторную задачу.
Ограничения исследования
Потенциальным ограничением в интерпретации наших результатов является трудность гарантировать четкое разделение между моторным обучением и моторным исполнением с помощью такой задачи, как FTT, в которой участники многократно воспроизводят как можно быстрее в течение 30 секунд за блок предопределенные, явно известные последовательности движений пальцами. Действительно, если участникам предлагается в качестве контрольного условия выполнить «более простую» базовую последовательную задачу постукивания (например, 1–2–3–4–1–2–3–4–...), существует еще последовательный компонент, который необходимо оптимизировать. Если, с другой стороны, им предлагается выполнить постукивание одним пальцем (или всеми пальцами одновременно), компоненты моторного движения соартикуляции полностью теряются, и эти две задачи различаются более чем на компонент моторного (последовательного) обучения. Теоретически, можно было бы оценить влияние обучения последовательности помимо моторного компонента, попросив участников изучить вторую, отличную последовательность и вычислить эффекты проактивной интерференции первой последовательности на приобретение второй, как это было сделано для задач моторной адаптации⁵⁰, но это не было реализовано в текущем протоколе. Чтобы хотя бы частично решить эту проблему, мы рассчитали индекс базовой производительности (BL) как среднее значение первых блоков FTT сессии обучения, предполагая, что на этом этапе участники еще не выучили последовательность, что делает ее потенциальной ситуацией моторного контроля. Как сообщалось выше, наши результаты не только выявили высокие коэффициенты корреляции между значениями поведения BMP и BL, но и то, что корреляционные анализы между параметрами СММ и оценками BL в основном дали аналогичные паттерны, как и с BMP (см. Дополнительную информацию, Таблица 6), поддерживая наше предположение о том, что корреляции между производительностью и параметрами состояния в значительной степени обусловлены индивидуальной моторной способностью, а не последовательным обучением, даже если мы признаем, что мы не можем полностью исключить вклад последнего.
Выбор количества состояний
Кроме того, было показано, что изменение числа состояний в анализах СММ может изменять характеристики всплесков каждого состояния⁵¹. Следовательно, выбор количества состояний (который должен быть предопределен в СММ) может повлиять на наблюдаемые результаты. Мы выбрали здесь априорное число восемь состояний, как на основе существующей литературы, так и на основании опыта нашей собственной лаборатории. Определение оптимального количества состояний важно как для избежания избыточной информации, так и для не упущения интересующей активности. Действительно, слишком низкое число состояний (например, 3) может привести к упущению потенциального разделения между активностью, связанной с моторным обучением и консолидацией памяти. С другой стороны, слишком большое число состояний может разделить моторную активность на несколько, более или менее избыточных состояний с низкими временными характеристиками. Например, параметр среднего времени жизни будет короче в этих состояниях, поскольку он распределен по нескольким состояниям, как показано Seedat et al.⁵¹. В основополагающей статье Бейкера и др.¹⁶ по СММ огибающих, авторы предположили, что оптимальное количество состояний может быть оценено с помощью модели с наименьшим значением свободной энергии, и в итоге выбрали 8 состояний как хороший компромисс между богатством и избыточностью. Кроме того, в недавней статье нашей группы⁵² число состояний было сокращено до 6, а топографии состояний были весьма схожи с теми, которые были получены с 8 состояниями²⁰‚³⁹, предполагая, что объем информации, полученный для 6 и 8 состояний, относительно схож. Хотя мы не можем исключить потенциальные ограничения, связанные с нашим априорным выбором использования 8 состояний, мы решили использовать стандартное априорное число, а не проводить параллельные анализы, манипулируя числом состояний, что привело бы нас к проблеме выбора лучших параметров анализа на основе их результатов, что является цикличным.
Заключение
Таким образом, наши результаты предполагают устойчивую, устойчивую связь между меж индивидуальными различиями в способности к моторной производительности и быстрой переходной нейронной динамикой в состоянии покоя. Отсутствие ассоциаций с поведенческими параметрами обучения и консолидации памяти, связанными с нейронной пластичностью, предполагает, что быстрая переходная динамика СП может не модулироваться опытом обучения и отражать нейронные пластические механизмы, по крайней мере, на коротком временном масштабе консолидации в рамках данного эксперимента.
Методы
Участники и процедура
Для этого эксперимента было привлечено тридцать четыре участника, согласно расчетной численности выборки (средний размер эффекта в внутрисубъектном дизайне с использованием двустороннего t-теста, с размером эффекта 0,5, уровнем значимости 0,05 и мощностью 0,8). Все испытуемые были молодыми и здоровыми правшами, не музыкантами и не профессиональными наборщиками, которые дали письменное информированное согласие на участие в данном исследовании, одобренном Этическим комитетом CUB Hôpital Erasme (№: P2016/553; CCB: B406201630539). Однако 7 участников пришлось исключить после предварительного анализа: 3 набора данных были отброшены из-за плохой моторной производительности; 1 набор данных имел поврежденный сигнал МЭГ, и 3 участника были идентифицированы как выбросы в анализе скрытой марковской модели из-за аберрантных, крайних значений. В общей сложности сообщаются данные 27 участников (11 женщин; средний возраст = 23,4 ± 2,7 лет, диапазон 18–29). Все методы выполнялись в соответствии с соответствующими руководящими принципами и нормами. Женщины-участницы тестировались во вторую неделю менструального цикла, чтобы избежать гормонального влияния на способности к моторному обучению⁵³. Напитки, содержащие кофеин, газированные напитки и любые другие стимуляторы были запрещены за 12 часов до тестирования.
По прибытии в лабораторию участники были подготовлены к записи МЭГ. Затем они прошли в МЭГ-сканере первую 5-минутную сессию состояния покоя (СП) (до обучения) в сидячем положении с открытыми глазами, сфокусированными на фиксирующем кресте на стене. Затем участники заполнили визуальные аналоговые шкалы (VAS) утомляемости и сонливости, чтобы выявить потенциальные эффекты сонливости и утомления. Сразу после этого они прошли обучение на 5-элементной задаче постукивания пальцами (FTT4; адаптированной из⁵⁴). В этой задаче (Рис. 1A) каждый палец соответствовал одной цифре (от 1 = мизинец до 4 = указательный палец), и участникам было поручено непрерывно воспроизводить 5-элементную последовательность движений пальцами (4–1–3–2–4) как можно быстрее и точнее ненатуральной рукой в течение 30 секунд (1 блок). Последовательность для воспроизведения постоянно отображалась на экране компьютера во время выполнения. Участники ознакомились с задачей в течение 2 демонстрационных блоков FTT, а затем прошли экспериментальную сессию обучения (LS) с двадцатью 30-секундными блоками FTT с 20-секундными перерывами между блоками, находясь в МЭГ-сканере. Сразу после задания участникам было разрешено покинуть кресло сканера и перемещаться по комнате. По окончании 20-минутного перерыва они заполнили VAS и провели вторую сессию МЭГ СП (после обучения). Наконец, прирост производительности с конца обучения оценивался путем выполнения двух блоков FTT (Тестовая сессия; Рис. 1B).
Поведенческие индексы моторной производительности и обучения
Производительность FTT оценивалась для каждого блока путем вычисления Индекса Глобальной Производительности (GPI), который учитывает как скорость, так и точность³¹. На основе показателей GPI мы определили лучшую моторную производительность (BMP), достигнутую каждым субъектом, как среднее значение двух блоков LS с наивысшими показателями GPI, что отражает вклад как моторики, так и последовательности. Мы также оценили базовую производительность (BL) как среднее значение блоков FTT 2 и 3 сессии обучения (блок 1 не учитывался в этом анализе, так как из-за начальной адаптации к задаче вариабельность была высокой с частыми остановками и нарушениями в выполнении последовательности), предполагая, что на этом этапе участники еще не выучили последовательность, что делает ее потенциальной ситуацией моторного контроля. Для измерения эволюции производительности во время сессии обучения мы вычислили Индекс Обучения (LI) как процентное изменение показателей GPI от BL до BMP. Наконец, мы оценили офлайн изменения производительности от конца обучения до тестовой сессии (т. е. эффект «буста») как процентное изменение от BMP до наилучшего показателя GPI, достигнутого после тестирования после обучения.
Приобретение данных нейровизуализации
Данные МЭГ записывались с использованием 306-канальной системы МЭГ полного охвата (Triux, MEGIN, Хельсинки, Финляндия), расположенной внутри легкой магнитно-экранированной комнаты (Maxshield, MEGIN, Хельсинки, Финляндия) в CUB Hôpital Erasme (Брюссель, Бельгия). Положение головы участников непрерывно отслеживалось в МЭГ-шлеме четырьмя катушками отслеживания головы. Положение катушек и около 300 точек головы определялось по анатомическим ориентирам с помощью электромагнитного трекера (Fastrak, Polhemus, Colchester, Vermont, USA). Мы применяли онлайн аналоговый полосовой фильтр в диапазоне 0,1–330 Гц для всех записей и оцифровывали сигнал с частотой дискретизации 1 кГц. Высокоразрешающие 3D T1-взвешенные церебральные магнитно-резонансные изображения (МРТ) участников были получены после записи МЭГ на 1,5 Т МРТ-сканере (Intera, Philips, Нидерланды).
Предварительная обработка данных
Метод разделения временного пространства сигналов⁵⁵ применялся в автономном режиме к непрерывным данным МЭГ для минимизации внешних магнитных помех и коррекции движений головы (Maxfilter v2.1, MEGIN, Финляндия). Затем данные фильтровались (автономный полосовой фильтр: 0,1–45 Гц), и применялся анализ независимых компонент (алгоритм FastICA с редукцией размерности до 30 компонентов, функция нелинейности гиперболического тангенса)⁵⁶ для визуальной инспекции. Независимые компоненты, соответствующие сердечным, глазным и системным артефактам, были отвергнуты путем регрессии их временной последовательности из данных полного ранга. Для проведения реконструкции источника, модели МЭГ-источника оценивались на основе 3D T1-взвешенной церебральной МРТ участников, анатомически сегментированной с использованием программного обеспечения FreeSurfer (версия 6.0; Martinos Center for Biomedical Imaging, Массачусетс, США). Системы координат МЭГ и МРТ были корегистрированы через три анатомические ориентировочные точки (носовая кость и уши) для первичной оценки положения головы и точки поверхности головы для ручной доработки (MRIlab, MEGIN Data Analysis Package 3.4.4, MEGIN, Хельсинки, Финляндия). Волюметрическая и регулярная сетка источников 5 мм была построена в шаблоне МРТ Монреальского неврологического института (MNI) и нелинейно деформирована на МРТ каждого участника с помощью программного обеспечения Statistical Parametric Mapping (SPM12, Wellcome Centre for Neuroimaging, Лондон, Великобритания). Наконец, 3D модель МЭГ-источника, связанная с этим пространством источников, оценивалась с использованием однослойного метода граничных элементов, реализованного в пакете MNEC.
Проекция источника данных МЭГ затем основывалась на оценке минимума нормы (MNE)⁵⁷. Матрица ковариации шума оценивалась из 5-минутных записей МЭГ в пустой комнате, пространственно фильтрованных методом разделения пространства сигналов⁵⁵ и временно отфильтрованных между 0,1 и 45 Гц. Параметр регуляризации MNE был фиксирован с использованием условия согласованности, выведенного в⁵⁸. Трехмерные временные ряды диполей проецировались на направление их максимальной дисперсии, и их сигнал огибающей сигнала извлекался с помощью преобразования Гильберта.
Динамический анализ скрытой марковской модели
Анализ проводился по конвейеру, описанному¹⁶‚³⁵, реализованному в GLEAN (https://github.com/OHBAanalysis/GLEAN). Основное отличие в нашей реализации заключается в том, что MNE используется как обратная модель, а не как формирователь луча (Beamformer), поскольку MNE позволяет исследовать состояния RSN, связанные с DMN, и, в частности, состояния, включающие задние срединные кортикальные области (т. е. предклинье и заднюю поясную кору)²⁰‚³⁹. Число переходных состояний было установлено равным 8 для соответствия предыдущим исследованиям СММ огибающих мощности МЭГ¹⁶‚²⁰‚³⁵‚³⁹. 8-состоятельная СММ выводилась из широкополосных (4–30 Гц) сигналов огибающих источников. Данные огибающих были понижены до 10 Гц с использованием скользящего среднего окна с 75% перекрытием (окна шириной 100 мс, скользящие каждые 25 мс), что привело к эффективному понижению до 40 Гц, центрированию, нормализации по глобальной дисперсии и временному объединению между участниками для разработки группового СММ анализа и между двумя сессиями СП для идентификации сетевых состояний, общих для пре- и пост-обучающих сессий (для дальнейшего обсуждения этой стратегии см., например,²⁰). Объединенные огибающие затем подвергались предварительному отбеливанию и редукции до 40 главных компонент. Наконец, алгоритм СММ³³‚⁵⁹ многократно запускался на этом наборе данных 10 раз (для учета различных начальных параметров и сохранения модели с наименьшей свободной энергией) для определения состояний, классифицирующих различные паттерны ковариации огибающей мощности. Алгоритм Витерби использовался для декодирования бинарных сигналов временно исключающей активации/деактивации состояния. На основе этих сигналов были оценены четыре параметра временной характеристики состояния: MLT (средняя продолжительность интервалов активного состояния), FO (общая доля времени активного состояния), MIL (средняя продолжительность интервалов неактивного состояния) и NO (общее количество посещений состояния). Эти индексы оценивались отдельно для каждого субъекта и сессии путем «разделения» временных рядов активации состояний. Карты мощности состояний были получены в результате частичной корреляции между временными рядами активации/деактивации состояний СММ и объединенными сигналами огибающей источников, что оценивает изменения мощности, специфичные для состояния, при активации состояния.
Статистические контрасты и корреляционные анализы с временными параметрами состояний СММ
Сравнение временных параметров состояний СММ до и после обучения оценивалось с использованием парного t-теста Уилкоксона. P-значения корректировались по Бонферрони для множественных сравнений с коэффициентом 7, т. е. количеством независимых состояний СММ (знание активации 7 из 8 состояний СММ полностью определяет активацию последнего состояния СММ из-за ограничения временного исключения, реализованного в алгоритме Витерби)²⁰. Для состояний СММ, показывающих значительный эффект пре- до пост-обучения, ранговые корреляционные анализы Спирмена исследовали взаимосвязь между их временными параметрами в пре- и пост-обучающих сессиях и поведенческими индексами. Непараметрические тесты были предпочтительны из-за более высокой устойчивости к выбросам, которые иногда возникают среди временных параметров состояний СММ, когда, например, один или несколько субъектов редко посещают одно состояние. Результаты считались значимыми при p < 0,05, скорректированными на множественные сравнения с коэффициентом 4, т. е. количеством рассматриваемых временных параметров СММ. Наконец, мы использовали байесовскую статистику для предоставления оценки вероятности нулевой гипотезы (тестирование значимости нулевой гипотезы; NHST) в случае незначительных результатов (результаты представлены в качестве дополнительного материала).