
Интересное сегодня

Введение
Для идентификации вида как культурного исследователи часто прибегают к анализу распределения поведенческих и/или артефактных форм (далее — поведений) в различных группах одного вида, используя так называемый метод исключения: если поведение присутствует в группе А, но отсутствует (или его частота крайне низка) в группе B, и если этот паттерн вряд ли может быть объяснён генетическими и/или экологическими различиями, то такое поведение следует считать находящимся под влиянием социального обучения. Это означает, что вид обладает культурой. Мы называем это распределительным подходом к культуре.
Распределительный подход и его ограничения
Распределительный подход лежит в основе нескольких исследований диких приматов, включая шимпанзе, орангутанов, паукообразных обезьян, горилл и бонобо, и он также использовался у неприматов. Исследования людей и доисторических гоминин также приняли, хотя иногда и неявно, распределительный подход к идентификации культуры. Хотя сейчас существуют дополнительные методы, направленные на выявление социального обучения в дикой природе, в этой статье мы сосредоточимся на классическом методе: методе исключения.
Социальное обучение и механизмы копирования
Хотя именно социальное обучение приводит к культуре, разные механизмы социального обучения приводят к разным типам культур. Многие исследователи поэтому дополнительно стремятся установить, использует ли вид конкретные механизмы социального обучения — как показатели различных типов культур. Эти исследователи часто (хотя и не всегда) сосредотачивались на обнаружении механизмов социального обучения, способных передавать specifics поведенческих и/или артефактных форм. Здесь социальное обучение гораздо более определено — оно должно быть социальным обучением, которое копирует specifics форм самого поведения (или ноу-хау, лежащего в основе поведения). Далее мы будем определять механизмы социального обучения, которые копируют specifics форм, как копирующее социальное обучение, или — для краткости — как «копирование».
Моделирование и результаты
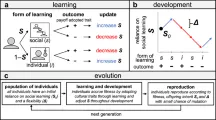
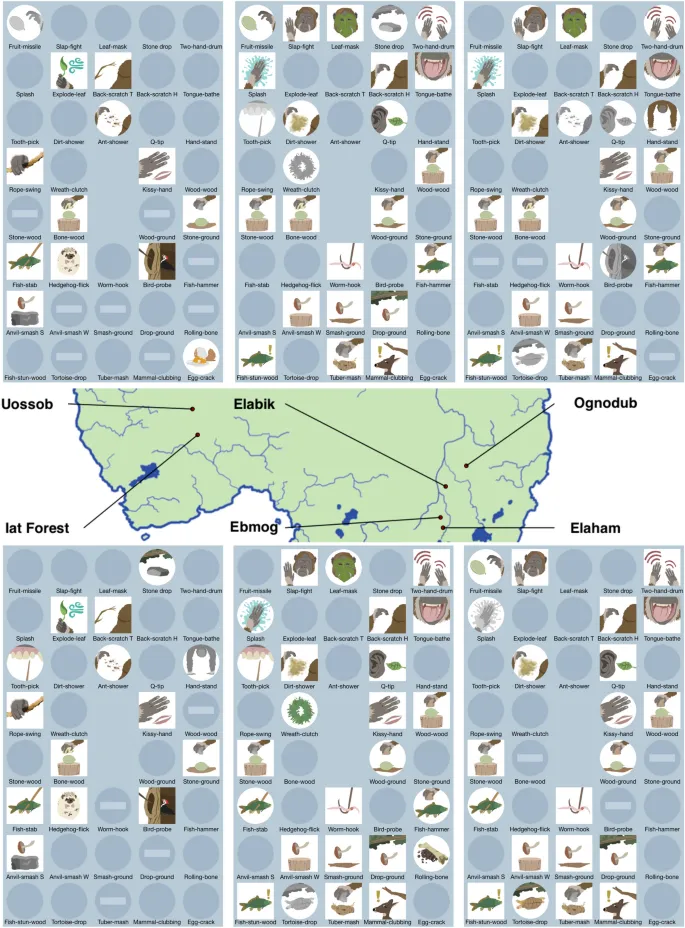
Мы построили индивидуализированную модель, воспроизводящую мир, населённый шестью популяциями «оранзи» — гипотетического вида приматов. Модель пространственно явная: популяции оранзи расположены в относительных позициях, аналогичных шести участкам шимпанзе в исследовании Уайтена и др. Это важно для определения потенциальных генетических предрасположенностей и экологической доступности, связанных с их возможными поведенческими или артефактными формами, или «поведениями». Размеры популяций также взяты из участков в Уайтене и др.
Навыки и ограничения оранзи
Оранзи подвержены упрощённому зависящему от возраста процессу рождения/смерти, вдохновлённому описаниями в Хилле и др. Шаг времени t симуляции представляет месяц жизни оранзи. Начиная с 25 лет (t = 300), есть 1% вероятность, что оранзи умрёт каждый месяц (максимальная продолжительность жизни ограничена 60 годами, т.е. t = 720). Число особей в популяции фиксировано, поэтому каждый раз, когда оранзи умирает, он заменяется новорождённым.
Поведения и состояние оранзи
В мире оранзи латентно возможны 64 поведенческие формы (свободно смоделированные на основе 65 поведений, закодированных в Уайтене и др., но сделанные чётным числом для удобства моделирования). Поведения разделены на две категории: 32 социальных и 32 связанных с едой поведения. Эти цифры были выбраны, чтобы напоминать категории поведений, рассмотренные в Уайтене и др. Поведения служат оранзи для выполнения различных биологических функций. Оранзи имеют «состояние», которое основано на том, сколько функций выполнено в двух основных категориях социальных и связанных с едой поведений.
Некопирующее социальное обучение
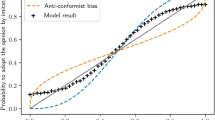
На каждом шаге времени все оранзи имеют вероятность — рассчитанную как описано выше — для индивидуального приобретения как социальных, так и связанных с едой поведенческих форм. Но какого именно поведения? Конкретная поведенческая форма, которую приобретёт оранзи, зависит как от частоты поведений, уже присутствующих в популяции, так и от экологической доступности и генетической склонности, связанной с поведенческой формой. Дополнительный параметр модели, S, контролирует вероятность того, что каждое приобретение социально опосредовано/запущено. Когда приобретение социально опосредовано/запущено, вероятность инновации каждого поведения Bi взвешивается по его пропорциональным случаям в популяции среди всех поведений той же категории (32 социальных поведения или 32 связанных с едой поведения). То есть, частота поведенческих форм может социально «катализировать» больше индивидуальных инноваций того же поведения: common поведения более вероятно индивидуально реинноваются на основе этого социального обучения. Когда инновация не социально опосредована/запущена, вероятность инновации каждого поведения случайна. Во всех случаях только одно поведение на особь может быть инновационно на каждом шаге времени.
Генетическая склонность и экологическая доступность
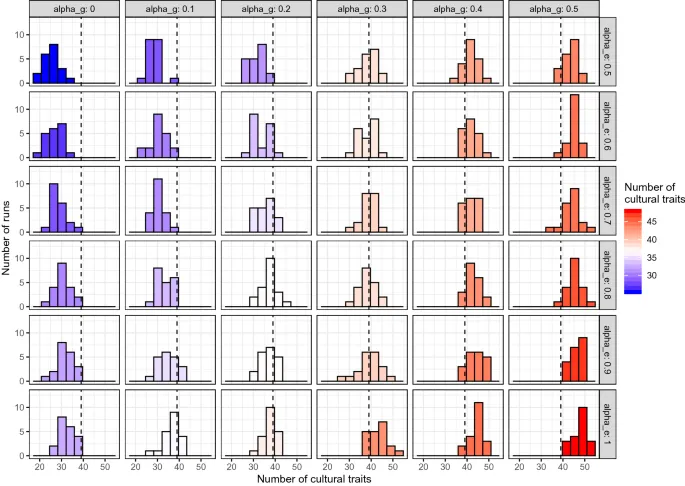
>Наконец, поведение, выбранное через описанную выше процедуру, фактически инновационно или нет в соответствии с его генетической склонностью и, в случае связанных с едой поведений, экологической доступностью. Генетическая склонность — это вероятность pg (0,1), назначенная независимо для каждого из 64 поведений (и не to be confused с инстинктами). Параметр модели, αg, определяет вероятность того, что генетическая склонность каждого поведения одинакова для всех шести популяций или различна. Если вероятность одинакова, pg случайно выбирается из равномерного распределения между 0 и 1. Если она различна, мы назначаем склонность, используя географический градиент. Мы выбираем случайную точку и вычисляем её расстояние до каждой популяции. Расстояния затем преобразуются в pg путём масштабирования их между 0 и 1, так что для самого дальнего участка, где pg = 0, связанное поведение не может быть выражено (см. Дополнительную информацию). Заметьте, что αg = 0 не означает, что нет генетических влияний на поведение, но что нет различий между популяциями в этом аспекте. Экологическая доступность — аналогично, вероятность pe (0,1), которая представляет likelihood находки ресурса или его nutritional value на каждом участке. Экологическая доступность назначается только связанным с едой поведениям и вычисляется так же, как pg, используя параметр αe чтобы определить вероятность экологической доступности being different or equal в шести популяциях.Обсуждение
Результаты нашей модели показывают, во-первых, как генетическая склонность и экологическая доступность влияют на formation культурных паттернов. Интуитивно, чем ниже генетическое и экологическое разнообразие, тем lower число черт, классифицируемых как «культурные» согласно распределительному методу. Заметьте, однако, что даже при полном отсутствии какого-либо экологического и генетического variation, т.е. с αe = 0 и αg = 0, некоторые культурные черты occur, согласно распределительному методу. Учитывая, что этот тип исключения невозможен в реальной жизни, этот outcome нашей модели somewhat reinforces валидность метода исключения как метода обнаружения культуры в более общем смысле (т.е. при отсутствии дальнейших inference regarding underlying learning mechanisms).
Выводы и значение
Наше моделирование не показывает и не может показать, что механизмы копирования не вызывают некоторые или даже все культурные распределения, наблюдаемые у диких приматов. Тесты этого конкретного утверждения требуют других методов (которые, однако, все указывают away от underlying копирования; см. введение) также см.52. Наш главный finding instead — replicating verbal claims, сделанные до нас — что такие паттерны не necessitate механизмы копирования. Поэтому эти паттерны (всё ещё) не могут быть использованы для inference копирования — другие, некопирующие механизмы социального обучения proved sufficient чтобы генерировать такие distributions в нашей модели. Мы надеемся, что мы further illustrated — здесь с помощью модели — что observation таких паттернов (распределительный подход) не может сам по себе suffice для infering существования механизмов копирования форм beyond индивидуального reach. В summary, inference копирования remains invalid — despite его continuing use в литературе (см. введение).





