Интересное сегодня
Способность предвидеть действия других является фундаментальной для социального взаимодействия, обеспечивая координацию, выявление намерений и адаптивное реагирование в динамичных ситуациях. Эффективное предвидение интегрирует предварительные ожидания и кинематические сигналы, извлекаемые из наблюдаемых движений (Gredin et al., 2023; Kemmerer, 2021). Этот интегративный процесс улучшает межличностную координацию и принятие решений в различных сферах, таких как спорт и общение. Однако процессы предвидения в реальных условиях редко протекают в расслабленных или неспешных временных рамках. Вместо этого люди часто сталкиваются со значительными временными ограничениями, требующими быстрого прогнозирования и принятия решений на основе неполной или неоднозначной информации. Поэтому необходимо исследовать, как временные ограничения влияют на интеграцию предварительных ожиданий и кинематических сигналов во время предвидения действий, и как эта модуляция отражается как в поведенческой производительности, так и в лежащих в основе нейронных процессах.
Теоретические основы
Байесовские модели предполагают, что мозг комбинирует предварительные ожидания с сенсорным вводом для генерации вероятностных прогнозов (Harris et al., 2023; Kilner et al., 2007; Knill & Pouget, 2004). При предвидении действий предварительная информация — такая как стратегические тенденции — предоставляет контекстуальные подсказки, в то время как кинематические характеристики, такие как поза и движение, предлагают доказательства в реальном времени (ElSourani et al., 2018; Koul et al., 2019). Оптимальные прогнозы требуют динамического взвешивания этих источников в зависимости от их надежности. Когда сенсорные сигналы неоднозначны, предварительные ожидания приобретают большее влияние; когда сенсорные доказательства ясны, они доминируют (Kelly & O’Connell, 2013).
Спортсмены особенно искусны в использовании упреждающих сигналов и поддержании производительности в условиях временных ограничений (Mann et al., 2007), предполагая, что некоторые люди могут лучше сохранять предвидительную функцию, несмотря на внешние временные требования. Однако байесовская интеграция предполагает достаточное время и ресурсы для обработки, которые могут быть нарушены при ограничениях (Loffing & CañalBruland, 2017). Ограниченное время может помешать детальному кинематическому анализу, смещая зависимость к эвристикам или упрощенным стратегиям (Tenenbaum & Land, 2009). Такие ограничения могут исказить процесс интеграции, приводя к субоптимальному взвешиванию предварительных и сенсорных сигналов и ухудшая качество предвидимого поведения. Основываясь на этой модели, мы предполагаем, что временные ограничения модулируют процесс интеграции, увеличивая зависимость от предварительной информации при одновременном снижении чувствительности к кинематическим деталям. Ожидается, что эта модуляция интеграции информации проявится как в поведенческой производительности, так и в нейронных индексах предвидимой обработки.
Стратегии обработки информации при временных ограничениях
Хотя временные ограничения могут нарушить оптимальную интеграцию, когнитивные системы адаптируются, перераспределяя ресурсы и отдавая приоритет низкозатратной информации, а не полностью отказываясь от вводов (Lavie, 2010; Shenhav et al., 2013). В условиях таких ограничений люди делают стратегические корректировки для поддержания эффективности и точности прогнозирования, часто путем смещения зависимости или модификации времени обработки. Временные ограничения, вероятно, упрощают интеграцию, отдавая предпочтение информации, требующей меньших когнитивных усилий. Предварительные ожидания, действующие через механизмы «сверху вниз» (top-down), предлагают эффективный путь для руководства прогнозами, ограничивая диапазон возможных исходов и снижая нагрузку на анализ сенсорных данных в реальном времени (Friston, 2010; Summerfield & Egner, 2009). В отличие от этого, обработка динамических кинематических сигналов требует более ресурсоемких механизмов «снизу вверх» (bottom-up). Таким образом, при временных ограничениях люди могут ускорить использование предварительной информации для поддержки предвидения, не отказываясь полностью от сенсорных доказательств.
Нейронные корреляты предвидения
Ожидается, что этот стратегический сдвиг проявится на нейронном уровне как усиленная предварительная активность после представления предварительного сигнала. Два взаимодополняющих нейронных маркера могут индексировать эту предвидимую вовлеченность. Контингентная отрицательная вариация (CNV, Contingent Negative Variation) — медленный корковый потенциал, возникающий между предупреждающим сигналом и побуждающим стимулом, — ассоциируется с ожиданием, распределением внимания и моторной подготовкой (Breska & Ivry, 2020; Hung et al., 2004; Wang et al., 2025). Увеличение амплитуды CNV в подготовительном интервале будет указывать на большую вовлеченность в предвидение, основанное на предварительных данных, в условиях временных ограничений. Дополнительно, альфа-колебания могут отражать обновление внутренних убеждений и контроль внимания в частотной области. Недавние исследования показывают, что более высокая альфа-активность кодирует более сильную силу байесовских убеждений и более эффективное распределение внимания (Haegens et al., 2011; Li et al., 2023). Таким образом, усиленная подготовительная альфа-активность может отражать когнитивный сдвиг в сторону стратегий предвидения, управляемых предварительной информацией, когда временные ограничения снижают способность к детальной сенсорной обработке.
Перцептивная обработка кинематики
Помимо подготовительной активности, временные ограничения могут также влиять на перцептивную обработку кинематических сигналов. Некоторые исследования предполагают, что при сжатых временных условиях люди больше полагаются на быструю перцептивную выборку для поддержания производительности (Cisek & Kalaska, 2010; Heitz, 2014). Данные айтрекинга (eyetracking) далее показывают, что эксперты демонстрируют более эффективные паттерны взгляда — более короткие фиксации и более ранние саккады, сфокусированные на цели — в условиях временных ограничений, что указывает на адаптивную реорганизацию визуального внимания (Vickers, 2007). Эти выводы поднимают возможность того, что временные ограничения могут улучшить извлечение или взвешивание кинематической информации. Однако нейрофизиологические исследования предполагают, что сенсомоторное моделирование — часто индексируемое подавлением мю-ритма — остается относительно стабильным и в большей степени формируется опытом, чем краткосрочными задачами (Del Percio et al., 2009; Denis et al., 2017; Fox et al., 2016). Эксперты последовательно демонстрируют более сильное подавление мю-ритма при наблюдении действий, направленных на цель, что отражает более надежные внутренние модели действий, тогда как новички демонстрируют слабые или отсутствующие реакции. Важно отметить, что такая модуляция мю-ритма оказывается относительно нечувствительной к контекстуальным переменным, таким как достоверность сигнала (Wang et al., 2025). Таким образом, в то время как исследования айтрекинга демонстрируют, как временные ограничения реорганизуют визуальное внимание в краткосрочной перспективе, подавление мю-ритма часто рассматривалось как маркер механизмов сенсомоторного моделирования, связанных с опытом, и относительно менее чувствительных к немедленным манипуляциям задачей. Эти выводы предполагают, что временные ограничения смещают стратегии предвидения к большей опоре на обработку «сверху вниз», при этом перцептивный анализ кинематических сигналов продолжает играть роль.
Цель исследования
Учитывая все вышесказанное, можно предположить, что временные ограничения могут влиять на предвидение действий, изменяя эффективность и время обработки информации, а не фундаментально изменяя перцептивное извлечение кинематических сигналов. В частности, временные ограничения могут вызывать более раннее вовлечение в предварительные ожидания и способствовать стратегическому упрощению интеграционных процессов, при этом оставляя основные механизмы сенсомоторного моделирования в значительной степени нетронутыми. Основываясь на этой модели, настоящее исследование изучило, как профессиональные баскетболисты адаптируют свою предвидимую обработку в условиях различных временных требований. Используя задачу предсказания, специфичную для спорта, мы объединили поведенческие и ЭЭГ-измерения для оценки того, поддерживает ли доменный опыт гибкое, контекстно-чувствительное предвидение. В этом мы стремимся прояснить, как мозг поддерживает адаптивное предвидение в быстро меняющихся, реальных условиях, и расширить существующую литературу, продемонстрировав, что временные ограничения изменяют интеграцию предварительной информации и кинематических сигналов у спортсменов-экспертов, тем самым продвигая наше понимание нейронной динамики предвидимой обработки.
Методы
Участники
В исследовании ЭЭГ приняли участие тридцать восемь профессиональных баскетболистов (средний возраст = 20,38 года, стандартное отклонение = 1,33; 19 женщин), все члены Китайской университетской баскетбольной ассоциации (CUBA). Поскольку CUBA является ведущим национальным студенческим соревнованием в Китае, все участники имели значительный соревновательный опыт в дополнение к своему тренировочному фону. Все тренировались ≥ 3 часа в день, 5 дней в неделю, более 8 лет и имели как минимум сертификат игрока Национального уровня второго класса. Ни у одного из участников не было истории алкогольной или наркотической зависимости или каких-либо неврологических или психических расстройств. Три участника были исключены из-за чрезмерных артефактов ЭЭГ, в результате чего окончательная выборка составила 35 правшей (средний возраст = 20,49, стандартное отклонение = 1,21; 17 женщин) с нормальным или скорректированным зрением. Все участники не были осведомлены о цели исследования и предоставили письменное информированное согласие. Исследование было одобрено этическим комитетом Шанхайского университета спорта (одобрение № 102772022RT069).
Стимулы
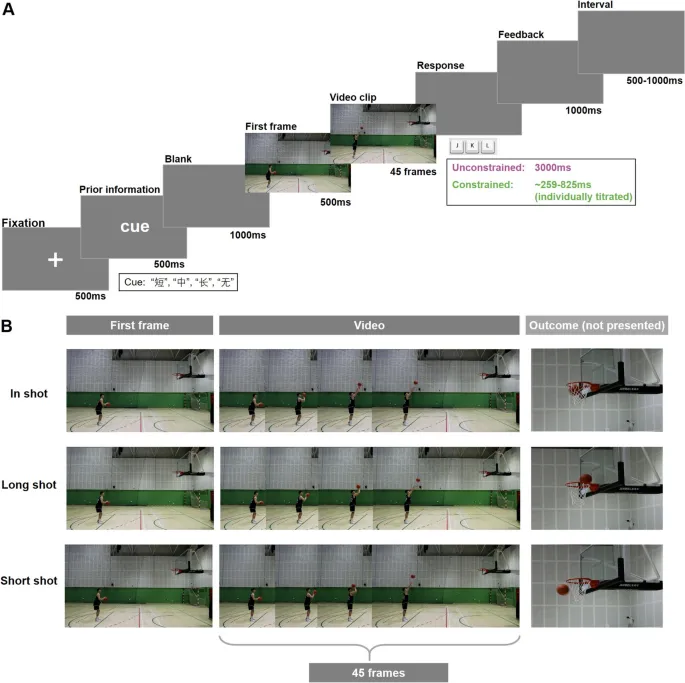
Стимулы состояли из специально записанных видеоклипов с окклюзией, снимающих двух профессиональных баскетболистов, выполняющих броски со штрафной линии (см. Рис. 1B). Видеозапись была сделана с частотой 60 Гц с помощью цифровой видеокамеры Canon EOS R6, расположенной на расстоянии 6 м от сагиттального вида. Два правши, один мужчина и одна женщина, выполнили три различных типа бросков: (1) попадание, когда мяч чисто попадал в корзину, не касаясь обода; (2) дальний бросок, когда мяч перелетал корзину; и (3) короткий бросок, когда мяч не долетал до корзины. Каждый игрок сделал 20 успешных попыток для каждого типа броска, чтобы обеспечить единообразие. Видеоклипы были отредактированы с использованием программного обеспечения Adobe Premiere для создания последовательностей окклюзии, охватывающих от 42 кадров до выпуска мяча до 2 кадров после выпуска. Примеры видеостимулов приведены в дополнительных материалах.
Рис. 1. Экспериментальный дизайн и стимулы. A. Последовательности испытаний задачи предвидения действий, специфичной для спорта. B. Три типа штрафных бросков, использованных в качестве стимулов: попадание, дальний бросок; и короткий бросок (примечание: исход броска не был представлен в стимулах).
Задача и процедура
Участники сидели примерно в 60 см от 17-дюймового монитора (разрешение 1024 × 768, частота обновления 60 Гц) в тускло освещенной, звукоизолированной комнате. Они располагали пальцами правой руки на клавишах «J», «K» и «L» для обозначения короткого, попадающего и дальнего броска соответственно. Задача включала два внутрисубъектных фактора: предварительная информация (присутствует против отсутствует) и временные ограничения (присутствуют против отсутствуют). В предварительном условии сигнал отображал один из трех китайских иероглифов — «短» (короткий), «中» (попадание) или «长» (дальний) — указывающий на вероятный исход предстоящего броска. В условии без предварительной информации сигнал отображал «无» (нет). Участникам было сообщено, что предварительные сигналы, как правило, верны, хотя точная точность не была указана. Чтобы стимулировать зависимость как от предварительной, так и от кинематической информации, конгруэнтность сигнала и исхода была установлена на уровне 60%, что значительно выше уровня 33,3% по шансам. Участникам было поручено предсказать исход броска на основе как предварительного сигнала (если он доступен), так и наблюдаемой кинематической информации. Эта процедура была адаптирована из нашей предыдущей работы (Chen et al., 2024; Wang et al., 2025), которая использовала уровень конгруэнтности 53,3%, что влияло на ответы спортсменов-экспертов, но обеспечивало ограниченную достоверность сигналов. Для повышения эффективности сигналов в настоящем исследовании уровень конгруэнтности был увеличен до 60%.
Каждое испытание начиналось с представления центрального крестика фиксации в течение 500 мс, за которым следовал сигнал (с предварительной информацией или без нее) в течение 500 мс и пустой экран в течение 1000 мс. Затем появлялся первый кадр видеоклипа в течение 500 мс, за которым следовал полный видеоклип. Участникам было поручено отвечать немедленно после окончания видео, чтобы минимизировать потенциальные артефакты ЭЭГ, связанные с движением. Обратная связь, указывающая «правильно», «неправильно» или «слишком медленно», отображалась в течение 1000 мс. В безусловном условии у участников было до 3000 мс для ответа. В обусловленном условии использовался индивидуальный крайний срок ответа, рассчитанный как медиана времени реакции каждого участника в безусловных испытаниях (диапазон: 259–825 мс, индивидуально подогнанный). Поздние ответы вызывали обратную связь «слишком медленно», и испытание немедленно заканчивалось. Межинтервальный интервал варьировался случайным образом от 500 до 1000 мс (см. Рис. 1A).
Эксперимент состоял из двух прогонов: одного безусловного и одного обусловленного. Каждый прогон включал четыре блока (два с предварительной информацией, два без нее) в дизайне ABBA. Каждый блок содержал 60 испытаний, с 1-минутными перерывами между блоками. Безусловный прогон всегда проводился первым, чтобы определить базовое время реакции каждого участника для установки крайнего срока в обусловленном прогоне. 5-минутный перерыв разделял два прогона, чтобы снизить утомляемость. Тот же набор из 60 видеоклипов случайным образом предъявлялся во всех блоках, и инструкции отображались в начале каждого. После каждого прогона участники заполняли NASA Task Load Index (NASATLX; Hart & Staveland, 1988) для оценки воспринимаемой когнитивной нагрузки. Перед основным экспериментом участники прошли 12 практических испытаний (по три на каждый тип сигнала), чтобы ознакомиться с процедурой.
Анализ поведенческих данных
Испытания с предварительной информацией были классифицированы как конгруэнтные (сигнал и исход совпадают) и инконгруэнтные (сигнал и исход не совпадают). Время реакции (RT) и точность рассчитывались для каждого участника по трем условиям: без предварительной информации, конгруэнтное и инконгруэнтное. Отдельные двухфакторные дисперсионные анализы (2 [Временные ограничения: безусловные, обусловленные] × 3 [Конгруэнтность: без предварительной информации, конгруэнтные, инконгруэнтные]) с повторными измерениями проводились для RT и точности. Следует отметить, что, поскольку участникам было поручено отвечать только после окончания видео, RT могут не отражать предвидимую обработку в реальном времени и анализировались исключительно для проверки манипуляции временными ограничениями. Точность служила основным показателем предвидимой производительности.
Чтобы оценить, как временные ограничения влияли на интеграцию информации, мы рассчитали разницу в точности между конгруэнтными и инконгруэнтными условиями предварительной информации при обоих временных режимах. Этот индекс зависимости от сигналов отражает опору на предварительную информацию, причем большие различия указывают на более сильное влияние сигналов. Для изучения согласованности стратегий интеграции в разных условиях были рассчитаны коэффициенты корреляции Пирсона для изучения взаимосвязи между зависимостью от сигналов в обусловленных и безусловных условиях. Наконец, парные t-тесты сравнивали оценки NASATLX по временным условиям, чтобы определить, отличалась ли воспринимаемая когнитивная нагрузка между безусловными и обусловленными условиями.
Регистрация и предварительная обработка ЭЭГ
Сигналы ЭЭГ записывались с помощью Brain Vision Recorder (версия 2.0; Brain Products GmbH, Германия) с 64 электродов AgCl, расположенных в соответствии с международной системой 10–20, с частотой дискретизации 1000 Гц. FCz служил онлайн-ссылкой, а AFz — заземлением. Вертикальные и горизонтальные электроокулограммы записывались с электродов, расположенных ниже левого глаза и на наружных спайках соответственно. Импедансы поддерживались ниже 5 кОм на протяжении всего процесса. Офлайн-предварительная обработка проводилась с использованием EEGLAB (Delorme & Makeig, 2004) и пользовательских скриптов MATLAB. Данные были повторно референцированы к среднему значению всех электродов и отфильтрованы с помощью фильтра нижних частот с конечной импульсной характеристикой (0,1–40 Гц). Был применен фильтр вырезания (48–52 Гц) для подавления сетевого шума 50 Гц. Анализ главных компонент (ICA) выполнялся для идентификации и удаления глазных артефактов, дополненный ручной проверкой. Затем данные были понижены до 500 Гц и сегментированы на 4,5-секундные эпохи (от −2 до +2,5 с относительно начала стимула, определяемого как начало первого кадра видеостимула). Коррекция базовой линии применялась с использованием 200-миллисекундного окна перед сигналом (от −1,7 до −1,5 с). Испытания, превышающие ±100 мкВ, были исключены, что привело к 14,5% отклонения. В среднем 108,5 (предварительная информация) и 103,5 (без предварительной информации) испытаний были сохранены в безусловном условии, и 101,2 (предварительная информация) и 96,8 (без предварительной информации) в обусловленном условии.
Анализ вызванных потенциалов (ERP)
Для оценки связанной с сигналом подготовительной нейронной активности до наблюдения за действием мы проанализировали CNV. Амплитуда CNV рассчитывалась как среднее напряжение в окне от −500 до 0 мс относительно начала первого кадра видео, на основе данных с коррекцией базовой линии. Этот анализ фокусировался на фронтоцентральных электродах (FC1, FCz и FC2; см. Рис. 3B), которые тесно связаны с обработкой, связанной с ожиданием (Kononowicz & van Rijn, 2014, Breska & Ivry, 2020). Для амплитуды CNV проводился двухфакторный дисперсионный анализ с повторными измерениями (2 [Временные ограничения: безусловные, обусловленные] × 2 [Предварительная информация: предварительная, без предварительной]), чтобы оценить влияние предварительной информации на подготовительную активность перед началом кинематических сигналов. Хотя испытания в условии предварительной информации включали как конгруэнтные, так и инконгруэнтные исходы, участники не могли определить конгруэнтность во время измерения. Поэтому амплитуда CNV сравнивалась между условиями с предварительной информацией и без нее, независимо от конгруэнтности.
Многомерная классификация паттернов
Многомерная классификация паттернов использовалась для декодирования сигналов ЭЭГ, связанных с предварительной информацией, в различных условиях временных ограничений (Bae & Luck, 2018; Wang et al., 2025). Эпохи охватывали от 200 мс до начала сигнала до 1500 мс после первого кадра видеоклипа. Для повышения эффективности декодирования данные ЭЭГ были понижены до 50 Гц (т.е. одна точка данных на 20 мс) и помечены по условию предварительной информации (предварительная против без предварительной). Все электроды служили признаками. Классификатор машины опорных векторов с кодами вывода, исправляющими ошибки (ECOC; Dietterich & Bakiri, 1994), был обучен с использованием функций fitcecoc() и predict() MATLAB. Пятикратная перекрестная проверка повторялась десять раз со случайными разделениями. Точность классификации усреднялась по фолдам и итерациям и сглаживалась скользящим средним из пяти точек. Для оценки того, превосходит ли точность декодирования уровень случайности (50%), проводился блочный тест перестановок с 10 000 итераций с использованием одновыборочных t-тестов по временным точкам. Кроме того, для сравнения точности декодирования между обусловленными и безусловными условиями проводился второй блочный тест перестановок с 10 000 итераций с использованием парных t-тестов по временным точкам. Множественные сравнения контролировались путем выявления значимых кластеров по временным точкам (для методологических деталей см. Bae & Luck, 2018).
Наконец, для связи декодирования с поведением, средняя точность декодирования по временным точкам выше уровня случайности извлекалась для каждого участника в каждом условии временных ограничений. Эти значения служили индивидуальными индексами нейронной чувствительности к предварительной информации. Затем рассчитывались корреляции Пирсона между этими значениями и поведенческим индексом зависимости от сигналов, анализируемые отдельно для обусловленных и безусловных условий.
Временно-частотный анализ
Данные ЭЭГ сегментировались от −2000 до 2000 мс относительно начала первого кадра видео. Временно-частотное разложение проводилось с использованием преобразования Фурье на коротких временах (STFT) с оконными функциями Ханнинга, оценивая мощность от 2–30 Гц с шагом 0,5 Гц. Событийно-связанные спектральные возмущения (ERSP) рассчитывались относительно базовой линии от −1700 до −1500 мс (предшествующей началу предварительного сигнала) и преобразовывались в децибелы (дБ) с использованием преобразования логарифмического отношения 10 × (Grandchamp & Delorme, 2011). Коррекция базовой линии применялась на уровне отдельных испытаний.
Мы сосредоточились на подготовительной альфа-активности (от −500 до 0 мс) и постстимульной мю-активности (от 500 до 2000 мс). Альфа-диапазон (8–13 Гц) анализировался на электродах PCz и Pz, областях, связанных с нисходящим контролем внимания и обработкой, связанной с ожиданием (Haegens et al., 2011, Li et al., 2023; см. Рис. 4B). Как и в анализе CNV, это окно предшествовало началу кинематического сигнала; следовательно, альфа-мощность анализировалась с использованием двухфакторного дисперсионного анализа с повторными измерениями (2 [Временные ограничения: безусловные, обусловленные] × 2 [Предварительная информация: предварительная, без предварительной]). Мю-мощность (8–13 Гц) анализировалась на C3 и C4, электродах над сенсомоторной корой, отражая моторное моделирование и предвидение действий (Fox et al., 2016, Denis et al., 2017; см. Рис. 5B). Мю-мощность анализировалась с использованием двухфакторного дисперсионного анализа с повторными измерениями (2 [Временные ограничения: безусловные, обусловленные] × 3 [Конгруэнтность: конгруэнтные, инконгруэнтные, без предварительной информации]). Во временно-частотных представлениях уменьшение мощности относительно базовой линии показано синим цветом, а увеличение — красным (см. Рис. 4A, 5A).
Для изучения взаимосвязи между нейронной колебательной активностью и производительностью предвидения действий корреляционные анализы были ограничены испытаниями без предварительной информации, поскольку предварительная информация не давала значимых основных эффектов. Этот подход изолировал влияние временных ограничений без конфаундинга от использования сигналов. Мы рассчитали ΔERSP (обусловленные–безусловные) для альфа- и мю-мощности и коррелировали их с соответствующими значениями ΔAccuracy. Кроме того, мы проверили, предсказывала ли ERSP-мощность в каждом временном условии точность предвидения действий, связывая колебательную динамику с производительностью.
Для всех дисперсионных анализов размеры эффекта сообщались как частичные эта-квадратные ($ta_{p}^{2}$) значения. Когда нарушения сферичности были указаны тестом Моучли, применялись коррекции Гринхауса-Гейссера для корректировки степеней свободы. Корректированные значения сообщаются повсеместно.
Результаты
Поведенческие данные
Субъективная рабочая нагрузка
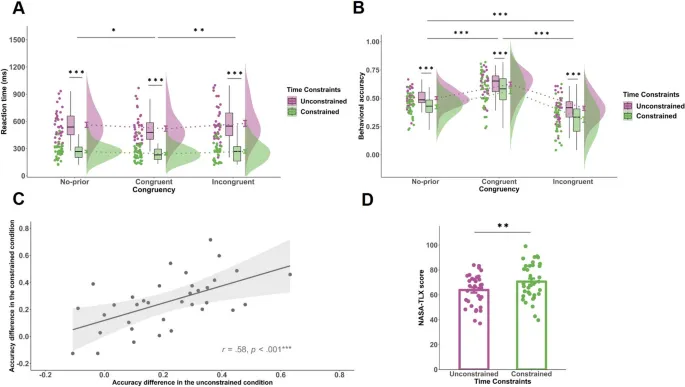
Оценки NASATLX были значительно выше в обусловленном условии, чем в безусловном, t(34) = 3,75, p = .001, d Коэна = 0,63, подтверждая, что временные ограничения увеличивали воспринимаемые требования к задаче (см. Рис. 2D).
Рис. 2. Поведенческие результаты. A. Среднее время реакции как функция временных ограничений и конгруэнтности. B. Поведенческая точность как функция временных ограничений и конгруэнтности. C. Корреляция зависимости от сигналов (разница в точности между конгруэнтными и инконгруэнтными испытаниями) по временным ограничениям. D. Оценки NASATLX в безусловных и обусловленных условиях. Уровни значимости обозначены как * p < .05, ** p < .01, *** p < .001.
Время реакции и точность
Основной эффект временных ограничений был значительным как для времени реакции (F(1, 34) = 61,14, p < .001, $ta_{p}^{2}$ = .64), так и для точности (F(1, 34) = 26,12, p < .001, $ta_{p}^{2}$ = .43). Время реакции было короче, а точность ниже в обусловленном условии.
Конгруэнтность сигнала также оказала значительное влияние на точность (F(2, 68) = 19,43, p < .001, $ta_{p}^{2}$ = .36) и время реакции (F(1,49, 67,78) = 4,81, p = .02, $ta_{p}^{2}$ = .12). Последующие пост-хок тесты показали, что конгруэнтные сигналы привели к более высокой точности по сравнению с инконгруэнтными и условиями без сигналов. Однако взаимодействие между временными ограничениями и конгруэнтностью не достигло статистической значимости ни для времени реакции (F(1,23, 68,72) = 1,21, p = .29, $ta_{p}^{2}$ = .03), ни для точности (F(2, 68) = 0,35, p = .71, $ta_{p}^{2}$ = .01). Это указывает на то, что влияние предварительной информации на предвидимую точность оставалось последовательным независимо от временных условий.
Зависимость от сигналов
Индекс зависимости от сигналов, рассчитанный как разница в точности между конгруэнтными и инконгруэнтными испытаниями, был сопоставим в обусловленных (M = .06, SD = .07) и безусловных (M = .07, SD = .08) условиях (t(34) = -0,78, p = .44, d = -0,13). Это подтверждает, что стратегическая опора на предварительную информацию не изменилась под воздействием временных ограничений.
Корреляция между зависимостью от сигналов в обусловленных и безусловных условиях была значительной (r = .65, p < .001), что указывает на стабильность индивидуальных стратегий интеграции информации (см. Рис. 2C).
Нейронные данные
Вызванный потенциал (CNV)
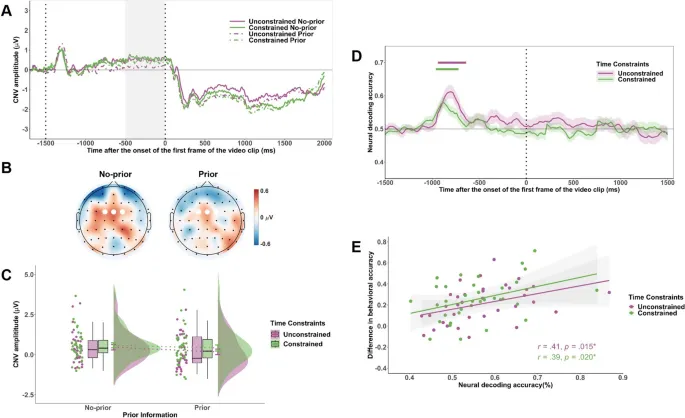
Анализ CNV выявил значимый основной эффект предварительной информации (F(1, 34) = 5,33, p = .027, $ta_{p}^{2}$ = .14), с большей амплитудой CNV в условии с предварительной информацией (M = -3,24 мкВ, SD = 2,55) по сравнению с условием без предварительной информации (M = -2,49 мкВ, SD = 2,31). Однако основные эффекты временных ограничений (F(1, 34) = 0,88, p = .35, $ta_{p}^{2}$ = .03) и взаимодействие между временными ограничениями и предварительной информацией (F(1, 34) = 0,09, p = .77, $ta_{p}^{2}$ = .003) не были значительными.
Многомерная классификация паттернов (MVPA)
MVPA показал, что сигналы ЭЭГ надежно различали условия с предварительной информацией и без нее в обоих временных условиях. Средняя точность классификации значительно превышала уровень случайности (50%) в течение длительных периодов времени после начала сигнала в обоих условиях. Сравнение между обусловленными и безусловными условиями не выявило значительных различий в точности декодирования (все p > .1).
Временно-частотный анализ: Альфа-ритм
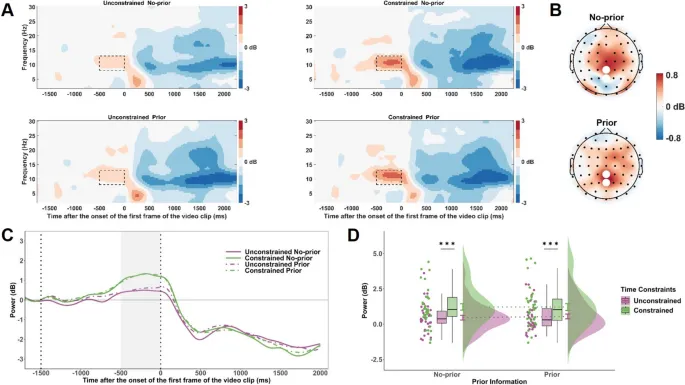
Во время подготовительного периода (от −500 до 0 мс до начала видео) наблюдалась повышенная синхронизация альфа-ритма (8–13 Гц) в парных электродах PCz и Pz в обусловленном условии по сравнению с безусловным (F(1, 34) = 6,75, p = .014, $ta_{p}^{2}$ = .17) (см. Рис. 4). Основной эффект предварительной информации (F(1, 34) = 0,95, p = .33, $ta_{p}^{2}$ = .03) и взаимодействие между временными ограничениями и предварительной информацией (F(1, 34) = 0,12, p = .73, $ta_{p}^{2}$ = .003) не были значительными.
Рис. 4. Временно-частотные результаты для активности в альфа-диапазоне во время подготовительного периода. A. Временно-частотные представления альфа-мощности (8–13 Гц) на электродах PCz и Pz для каждой комбинации условий временных ограничений и конгруэнтности. Черные пунктирные рамки обозначают окно анализа (от −500 до 0 мс) относительно начала видеостимула. B. Топографические карты разницы альфа-мощности между обусловленными и безусловными условиями (обусловленные минус безусловные), показанные отдельно для предварительных и непредварительных условий в окне времени от −500 до 0 мс. Белые точки представляют электроды PCz и Pz. C. Средние волновые формы альфа-мощности во времени на электродах PCz и Pz, построенные отдельно для каждой комбинации условий временных ограничений и конгруэнтности. Затененная серая область отмечает окно анализа (от −500 до 0 мс). D. Средняя альфа-мощность в окне от −500 до 0 мс по условиям временных ограничений и конгруэнтности. Уровни значимости обозначены как *** p < .001.
Временно-частотный анализ: Мю-ритм
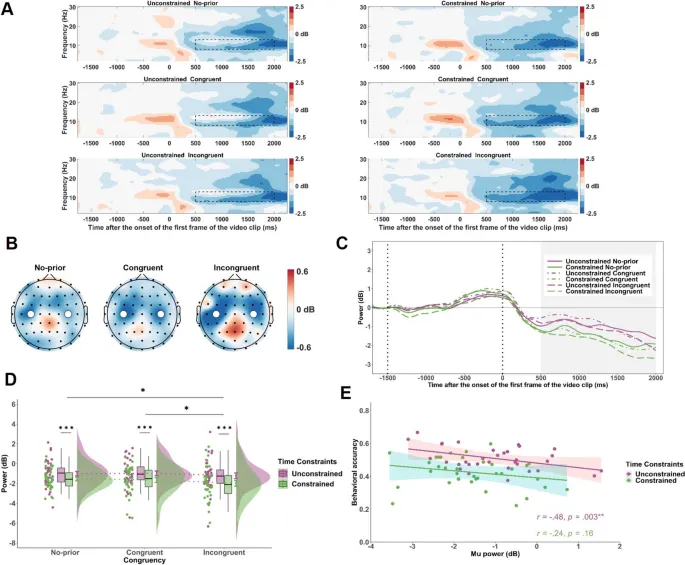
Во время перцептивной обработки кинематической информации (от 500 до 2000 мс после начала видео) наблюдалось более сильное подавление мю-ритма (8–13 Гц) в электродах C3 и C4 в обусловленном условии по сравнению с безусловным (F(1, 34) = 9,88, p = .003, $ta_{p}^{2}$ = .22) (см. Рис. 5). Отмечалось более сильное подавление мю-ритма в конгруэнтных и инконгруэнтных условиях по сравнению с условием без предварительной информации (F(2, 68) = 3,56, p = .034, $ta_{p}^{2}$ = .09).
Рис. 5. Временно-частотные результаты для активности в мю-диапазоне во время наблюдения за действием. A. Временно-частотные представления мю-мощности (8–13 Гц) на электродах C3 и C4 для каждой комбинации условий временных ограничений и конгруэнтности. Черные пунктирные рамки обозначают окно анализа (от 500 до 2000 мс) относительно начала видеостимула. B. Топографические карты разницы мю-мощности между обусловленными и безусловными условиями (обусловленные минус безусловные), показанные отдельно для конгруэнтных, инконгруэнтных и непредварительных условий в окне времени от 500 до 2000 мс. Белые точки представляют электроды C3 и C4. C. Средние волновые формы мю-мощности во времени на электродах C3 и C4, построенные отдельно для каждой комбинации условий временных ограничений и конгруэнтности. Затененная серая область отмечает окно анализа (от 500 до 2000 мс). D. Средняя мю-мощность в окне от 500 до 2000 мс по условиям временных ограничений и конгруэнтности. E. Корреляция между средним мю-ритмом (обусловленные минус безусловные) и разницей в точности (обусловленные минус безусловные) для предвидимой задачи. Уровни значимости обозначены как * p < .05, ** p < .01.
Связь между нейронной активностью и поведением
Корреляционный анализ выявил, что более сильное подавление мю-ритма в безусловном условии было связано с более высокой предвидимой точностью (r = .48, p = .003). Однако эта корреляция отсутствовала в обусловленном условии (r = .24, p = .16). Альфа-ритм не показал значимых корреляций с точностью ни в одном из условий.
Обсуждение
Данное исследование изучало, как временные ограничения влияют на интеграцию предварительных ожиданий и кинематической информации при предвидении действий у спортсменов-экспертов. В то время как временные ограничения значительно ухудшали точность предсказаний, поведенческая польза от конгруэнтных предварительных сигналов оставалась стабильной, предполагая сохраненную стратегическую опору на предварительную информацию. Многомерная классификация паттернов показала, что нейронные сигналы, предшествующие началу действия, надежно различали испытания с предварительной информацией и без нее в обоих условиях временных ограничений. Временные ограничения усиливали подготовительную активность альфа-ритма и увеличивали подавление мю-ритма во время кинематической обработки; однако только подавление мю-ритма в безусловном условии было связано с лучшей производительностью. В целом, результаты демонстрируют, что временные ограничения изменяют нейронную динамику предвидимой обработки, даже несмотря на сохраненную поведенческую опору на предварительные сигналы.
Снижение предвидимой точности под временными ограничениями
В соответствии с предыдущими исследованиями в области перцептивного принятия решений и моторной производительности (Kocher & Sutter, 2006; Vickers, 2007), наши выводы демонстрируют, что временные ограничения значительно ухудшают точность предвидения действий у спортсменов-экспертов. Хотя это снижение часто объясняется компромиссом между скоростью и точностью (Bogacz et al., 2010; Heitz, 2014), предвидимые решения в спорте также опираются на интеграцию предварительных ожиданий с разворачивающимися кинематическими сигналами (Abernethy et al., 2005; CañalBruland & Mann, 2015). Таким образом, временные ограничения могут нарушать производительность не только за счет сокращения времени ответа, но и за счет вмешательства в процессы восприятия, интеграции или принятия решений. Три потенциальных механизма могут объяснить наблюдаемое снижение производительности: (1) нарушение обработки кинематической информации, (2) измененная опора на предварительные ожидания и (3) нарушение интеграции предварительной и сенсорной информации. В последующих разделах мы оценим каждую возможность с помощью конвергентных данных из поведенческой производительности, субъективной рабочей нагрузки и нейронной динамики.
Перцептивная обработка и субъективная нагрузка
Одно из возможных объяснений снижения предвидимой точности в условиях временных ограничений — недостаточность обработки динамической кинематической информации. В поддержку этой точки зрения участники сообщали о более высокой субъективной рабочей нагрузке по NASATLX в условиях временных ограничений, указывая на то, что ограниченное время не только сокращало продолжительность обработки, но и увеличивало потребности в когнитивных ресурсах. В дополнение к этому, более сильная десинхронизация мю-ритма в условиях временных ограничений — обычно отражающая большую моторную симуляцию или вовлечение зеркальной системы — предполагает, что участники больше полагались на внутренние модели действий для компенсации сниженного восходящего ввода (Denis et al., 2017; Fox et al., 2016; Hari et al., 1998; Muthukumaraswamy & Johnson, 2004; Pineda, 2005). Однако эта повышенная сенсомоторная активация не улучшила производительность. Фактически, обычная связь между подавлением мю-ритма и лучшим предвидением, наблюдаемая в безусловном условии, исчезла при ограничениях. Это расхождение предполагает, что, хотя механизмы моторной симуляции были более активно вовлечены, обработка могла быть поверхностной, фрагментированной или плохо интегрированной. Одна из возможностей заключается в том, что активация сенсомоторных цепей отражает реакцию по умолчанию или компенсаторную реакцию на высокие требования задачи, а не оптимизированную стратегию обработки информации. Альтернативно, возросшая зависимость от моторной симуляции могла произойти за счет более глубокого перцептивного анализа или более адаптивной интеграции сигналов, что привело к увеличению субъективной нагрузки, но снижению предвидимой полезности. В любом случае, эти выводы подчеркивают, что при временных ограничениях большее нейронное вовлечение не обязательно приводит к лучшей обработке.
Стабильность опоры на предварительную информацию
Другое возможное объяснение заключается в том, что временные ограничения могут изменять саму стратегию принятия решений, в частности, снижая опору на предварительные ожидания. Однако эта интерпретация не подтверждается текущими данными. Поведенчески, участники последовательно демонстрировали лучшую производительность на конгруэнтных, чем на инконгруэнтных испытаниях, независимо от временного условия, и не было обнаружено взаимодействия между временными ограничениями и конгруэнтностью. Это указывает на то, что влияние предварительной информации на предвидимую точность оставалось стабильным. Электрофизиологические результаты далее подкрепляют эту интерпретацию. Предыдущие исследования показали, что в спортивных контекстах эффективные направляющие сигналы могут усиливать амплитуды CNV, отражая повышенное ожидание и моторную подготовку к ожидаемому действию (Hung et al., 2004; Wang et al., 2025). Однако в данном исследовании амплитуда CNV не различалась между условиями предварительного сигнала или временными ограничениями, что указывает на то, что подготовительное распределение сенсомоторных ресурсов не модулировалось временным давлением. В сочетании со стабильными поведенческими эффектами конгруэнтности эти выводы предполагают, что спортсмены продолжали полагаться на предварительную информацию в той же степени и с одинаковой подготовительной вовлеченностью, независимо от времени, доступного для ответа. Следовательно, снижение точности прогнозирования при временных ограничениях маловероятно связано с уменьшением использования предварительных знаний. Вместо этого, согласованность эффектов конгруэнтности и амплитуд CNV между условиями указывает на сохраненную предвидимую стратегию, что еще больше сужает вероятное место нарушения до более поздних стадий процесса интеграции.
Нейронные адаптации при временных ограничениях
Хотя MVPA показал надежное декодирование предварительных и непредварительных испытаний при обоих временных ограничениях, различия во времени начала декодирования не были статистически значимыми. Тем не менее, постоянное присутствие декодируемых сигналов и численные тенденции к более раннему вовлечению в обусловленных условиях предполагают, что спортсмены могут начинать обработку предварительных сигналов раньше, когда времени меньше. Эта интерпретация, хотя и предварительная, согласуется с взглядом на то, что временные ограничения вызывают предвидимые корректировки, направленные на компенсацию ограниченного доступа к кинематической информации.
Дополнительное подтверждение предвидимых корректировок при временных ограничениях дают наши выводы об альфа-диапазоне. В частности, мы наблюдали усиленную синхронизацию альфа-ритма в париетальных областях во время подготовительной фазы в обусловленном условии. Альфа-синхронизация обычно ассоциируется с нисходящим тормозным контролем, отражающим подавление релевантной информации и облегчение обработки, релевантной цели (Klimesch, 2012). В контексте предвидения действий, повышенная альфа-активность может указывать на сдвиг к более внутренне управляемому прогнозированию, позволяя мозгу готовиться к входящим кинематическим сигналам на основе предварительных ожиданий. Эта интерпретация согласуется с моделями предиктивного кодирования, которые предполагают, что мозг постоянно генерирует и обновляет внутренние модели для снижения сенсорной неопределенности (Friston, 2005; Smith et al., 2021). При временных ограничениях опора на такие предиктивные модели может стать более выраженной, обеспечивая более быстрые, но менее гибкие предвидимые ответы. Этот сдвиг может также объяснить наблюдаемое увеличение когнитивной нагрузки и подавления мю-ритма, поскольку когнитивные ресурсы перераспределяются для примирения предварительных ожиданий с ограниченной обработкой сенсорного ввода. В поддержку этой точки зрения, недавние исследования показали, что контекстуальные априорные данные могут формировать понимание действий, даже когда кинематическая информация искажена или неоднозначна (Amoruso et al., 2019; Bianco et al., 2024; Magnaguagno et al., 2022). Наши выводы расширяют это понимание, демонстрируя, что временные требования могут усиливать нисходящую модуляцию во время предвидения, что отражается в усиленной альфа-синхронизации. В целом, хотя поведенческая опора на предварительную информацию остается стабильной, временные ограничения, по-видимому, изменяют лежащую в основе нейронную стратегию, способствуя режиму обработки, более ориентированному на предсказание. Это подчеркивает способность мозга гибко адаптировать свои предвидимые механизмы для решения задач во временных ограниченных средах.
Ограничения исследования
Несколько ограничений данного исследования заслуживают внимания. Во-первых, манипуляция временными ограничениями была направлена на время выполнения ответа, но не ограничивала представление стимулов, оставляя неясным, приведет ли ограничение сенсорного воздействия к аналогичным эффектам предвидения. Во-вторых, предварительные сигналы предъявлялись с фиксированной достоверностью, что препятствовало оценке того, как временные ограничения влияют на гибкое взвешивание предварительной информации в зависимости от ее надежности. В-третьих, исследование включало только спортсменов-экспертов без группы новичков, что затрудняло определение того, являются ли наблюдаемые предвидимые корректировки специфичными для опыта или отражают более широкие когнитивные адаптации. Однако предыдущие работы нашей группы показали, что эксперты используют предварительную информацию более стабильно и избирательно, чем новички при перцептивном конфликте (Chen et al., 2024), предполагая, что опыт может помочь сохранить предвидимые стратегии, когда когнитивные ресурсы ограничены. Будущие исследования должны сравнивать спортсменов с разным уровнем опыта, чтобы прояснить, как опыт формирует временную гибкость в интеграции информации. Кроме того, включение комплементарных методологий могло бы предоставить конвергентные доказательства на различных уровнях анализа. Например, функциональная магнитно-резонансная томография (фМРТ) могла бы выявить механизмы крупномасштабной сети, поддерживающие предвидимую обработку при временных ограничениях, в то время как неинвазивные методы стимуляции мозга, такие как транскраниальная магнитная стимуляция (ТМС), могли бы установить причинные вклады моторных и париетальных областей в предвидение, основанное на предварительной информации. Наконец, безусловное условие всегда проводилось первым для установления базового времени реакции. Хотя этот подход позволил индивидуально калибровать временные ограничения, он также создал возможность для эффектов порядка, поскольку участники могли получить больше практики перед обусловленным условием. Тем не менее, обусловленное условие последовательно приводило к снижению точности прогнозирования и изменениям в нейронной динамике, что подтверждает интерпретацию того, что эти эффекты обусловлены временными требованиями, а не только практикой.
Заключение
Данное исследование демонстрирует, что временные ограничения ухудшают предвидение действий у спортсменов-экспертов, даже несмотря на то, что их поведенческое использование предварительной информации остается стабильным. Вместо нарушения предвидимых стратегий, временные требования, по-видимому, изменяют лежащие в основе нейронные процессы, на что указывают повышенная подготовительная активность альфа-ритма и более сильное подавление мю-ритма во время кинематической обработки. Эти результаты предполагают, что спортсмены адаптируются, смещаясь в сторону более внутренне управляемой, основанной на ожиданиях обработки в подготовке и вовлекая компенсаторное моторное моделирование во время наблюдения за действием. Эта нейронная корректировка отражает гибкую стратегию интеграции в условиях временных ограничений, предлагая представление о том, как мозг поддерживает предвидимую производительность в быстро меняющихся, неопределенных средах.