
Интересное сегодня


Исследование фокуса внимания как нового индикатора эмоционального состояния у животных
У людей различные эмоциональные состояния могут приводить к расширению или сужению фокуса внимания. Исследовано ли это явление у животных, пока не выяснено. Если это подтвердится, измерение фокуса внимания может стать новым когнитивным методом оценки благополучия животных. В данном исследовании мы поставили целью изучить тест фокуса внимания как меру эмоционального состояния у животных. Для этого мы индуцировали четыре предположительно различных эмоциональных состояния у собак (N = 10), различающихся по валентности (позитивная, негативная) и уровню возбуждения (высокий, низкий), в двух различных контекстах вознаграждения (пищевые вознаграждения в Эксперименте 1, социальные вознаграждения в Эксперименте 2), а затем оценили поведенческие реакции собак в тесте фокуса внимания. Мы также регистрировали параметры вариабельности сердечного ритма (ВСР) как дополнительные подтверждающие аффективные индикаторы.
Эксперимент 1: Пищевые вознаграждения
В Эксперименте 1 собаки продемонстрировали сужение фокуса внимания после индукции обоих позитивно окрашенных эмоциональных состояний. Тот факт, что собаки находились в позитивном состоянии, подтверждался снижением стандартного отклонения нормальных интервалов RR (SDNN) и снижением показателей низкой частоты (LF) и очень низкой частоты (VLF) ВСР.
Эксперимент 2: Социальные вознаграждения
В Эксперименте 2, при исследовании реакций на социальные вознаграждения, мы не выявили статистически значимых различий в фокусе внимания между эмоциональными состояниями, однако собаки демонстрировали несколько суженный фокус внимания в негативно окрашенных эмоциональных состояниях. Показатель LF имел тенденцию к снижению в условиях позитивного состояния с высоким уровнем возбуждения. В заключение, наше исследование предоставляет первое указание на то, что эмоциональные состояния могут изменять фокус внимания и у животных. Результаты оправдывают дальнейшие исследования этого подхода для использования в оценке благополучия животных, хотя для уточнения прогнозов необходимы дополнительные исследования.
Введение
У людей, а в последнее время и у животных, установлена сильная двунаправленная связь между когнитивными процессами и эмоциями1. Эмоции определяются как кратковременные ощущения, вызванные конкретным событием, и связаны с отчетливыми физиологическими, поведенческими и когнитивными реакциями2,3. Определенные эмоциональные состояния могут приводить как к сужению, так и к расширению внимания у людей4. Сужение внимания можно описать как «не видеть леса за деревьями», тогда как расширение внимания описывает «видение всего леса». Однако еще не исследовано, происходит ли подобное расширение и сужение фокуса внимания у животных при переживании ими различных эмоциональных состояний, и если да, то не может ли это служить новым когнитивным индикатором благополучия животных.
Фокус внимания может быть измерен с помощью теста на предпочтение глобальных/локальных элементов, который использует визуальные стимулы, состоящие из одного большого элемента, называемого глобальным, который, в свою очередь, построен из нескольких меньших элементов, называемых локальными. Широкий фокус внимания определяется как предпочтение глобальных элементов, тогда как узкий фокус внимания определяется как предпочтение локальных элементов. Тревожные люди быстрее обнаруживают локальные элементы в тесте на предпочтение глобальных/локальных элементов (т. е. у них узкая фокусировка внимания) по сравнению с нетревожными людьми5,6. В то время как люди в позитивном эмоциональном состоянии обычно демонстрируют расширение фокуса внимания7,8. Узкий фокус внимания гарантирует, что нерелевантные стимулы и восприятия могут быть проигнорированы, когда индивиды пытаются достичь цели (например, получить доступ к ресурсу или избежать опасности)4,8,9. Однако постоянное сужение внимания к одному типу стимула может затруднить переключение внимания на другие стимулы, которые могут быть более необходимы, или когда появляются новые непредвиденные стимулы10. С другой стороны, широкий фокус внимания, возникающий при переживании позитивных эмоций, связан с более открытой и целостной обработкой информации, что может способствовать обнаружению новой информации и исследованию новых возможностей11. Способность сужать и расширять фокус внимания в зависимости от текущего эмоционального состояния поэтому является существенной для адекватного реагирования на различные ситуации.
Помимо валентности эмоции, на фокус внимания у людей влияла мотивационная интенсивность4: позитивные эмоции возникают, когда люди мотивированы приближаться к вознаграждающим стимулам (например, возбуждение), но также и когда они успешно избегают негативных стимулов (например, облегчение). Негативные эмоции возникают, когда люди испытывают и/или предвидят вредные стимулы (например, страх) и когда позитивные стимулы удаляются (например, фрустрация)12. В литературе по благополучию животных эмоции часто описываются двухмерной моделью, состоящей из оси валентности (от позитивной к негативной) и оси возбуждения (от высокого к низкому)13. Концепция мотивационной интенсивности и возбуждения тесно связаны: эмоции, которые сильно мотивируют, часто также сильно возбуждают (например, ожидание пищевого вознаграждения), хотя в литературе по человеку есть предположения, что это не всегда так4,14. Тем не менее, для простоты, и поскольку в данной статье мы представляем исследование с животными, мы выбрали термин «возбуждение» вместо термина «мотивационная интенсивность».
Вкратце, фокус внимания отражает взаимодействие между валентностью эмоции и уровнем возбуждения, связанным с ней, тем самым нюансируя реакцию в конкретной ситуации. У людей было показано, что в то время как переживание достижения денежных выигрышей (позитивная эмоция с низким возбуждением) ассоциировалось с широким фокусом внимания, предвкушение денежных выигрышей (позитивная эмоция с высоким возбуждением) приводило к сужению фокуса внимания15. Более того, даже если фокус внимания индивидов сужался после просмотра изображения аппетитного десерта, он становился еще более узким при увеличении возбуждения, поскольку им давали понять, что они съедят десерт16. Относительно негативных эмоций, состояния с высоким возбуждением, такие как отвращение17 и тревога18, приводили к более узкому фокусу внимания, чем состояния с низким возбуждением, такие как грусть17.
Тест на предпочтение локальных/глобальных элементов уже был адаптирован для животных с целью сравнения когнитивной обработки у разных видов (например, 19,20). Однако эти исследования проводились без манипуляции эмоциональным состоянием. Большинство нечеловекообразных приматов имеют тенденцию к узкому фокусу внимания21,22,23. В то время как другие виды животных, такие как медоносные пчелы20, рыбы24 и птицы25,26, имеют локальное предпочтение (но см. 27 для исключения). Предпочтения собак в отношении локальной или глобальной информации варьировались между особями, что предполагает большую индивидуальную вариативность у собак19.
Для получения дополнительных мер эмоционального состояния, которые могли бы помочь в интерпретации поведенческих данных и валидации индукции различных эмоциональных состояний, мы также регистрировали параметры сердечного ритма (ЧСС) и вариабельности сердечного ритма (ВСР)28. Увеличение ЧСС обычно интерпретируется как увеличение возбуждения как у людей29, так и у собак30, хотя ЧСС также может указывать на валентность у людей31,32. Индексы ВСР часто интерпретируются как показатели валентного компонента эмоциональных состояний. Например, снижение стандартного отклонения нормальных интервалов RR (SDNN), среднеквадратичного отклонения последовательных разностей (RMSSD), показателей ВСР высокой частоты (HF) и низкой частоты (LF) было показано как признак позитивной валентности у собак30,33, в то время как увеличение RMSSD указывало на негативную валентность33, хотя см. 34.
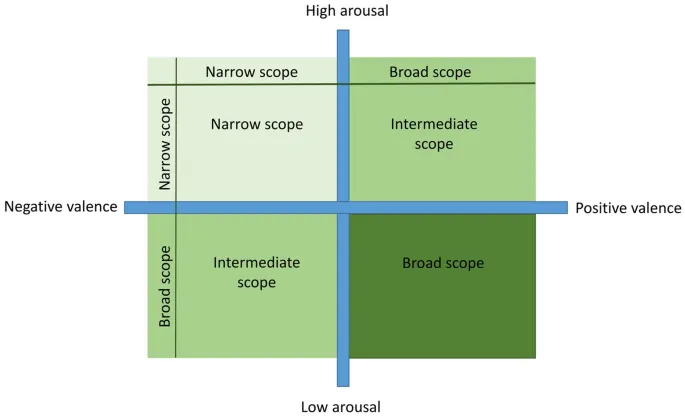
Цель данного исследования заключалась в изучении методологии оценки фокуса внимания как нового индикатора эмоционального состояния у животных. Мы использовали собак, поскольку они ранее показали себя как подходящая модель для исследования когнитивных и эмоциональных процессов (например, см. 19,35). Мы стремились индуцировать различные эмоциональные состояния, варьирующиеся по валентности (позитивная, негативная) и уровню возбуждения (высокий, низкий), в двух контекстах, известных как вознаграждающие для собак; пищевое вознаграждение и социальное взаимодействие30,36,37,38. Мы предположили, что позитивные эмоции в целом приведут к более широкому фокусу внимания по сравнению с негативными эмоциями. Кроме того, мы предположили, что возбуждение также повлияет на фокус внимания, причем состояния с высоким уровнем возбуждения приведут к более узкому фокусу внимания по сравнению с состояниями с низким уровнем возбуждения. Менее понятно, как они взаимодействуют. Таким образом, мы выдвигаем только два конкретных прогноза. А именно, что собаки в позитивном состоянии с низким уровнем возбуждения будут иметь самый широкий фокус внимания, а собаки в негативном состоянии с высоким уровнем возбуждения – самый узкий. Мы предполагаем, что собаки в позитивном состоянии с высоким уровнем возбуждения или в негативном состоянии с низким уровнем возбуждения должны быть промежуточными, но это будет зависеть от относительного вклада валентности и возбуждения (Рис. 1). В данном исследовании мы также измеряли индексы ВСР, предполагая, что ЧСС будет выше в состояниях с высоким возбуждением, а валентность эмоции будет связана с различными индексами ВСР, тем самым подтверждая индукцию эмоциональных состояний.
Материалы и методы
1. Субъекты
Мы оценили, что выборка из примерно 16 собак была бы достаточной для решения нашего исследовательского вопроса, согласно нашим первоначальным расчетам мощности (с 95% доверительным интервалом и 80% мощностью) на основе пилотного исследования фокуса внимания. Поэтому исследование началось с 21 лабораторной собаки породы бигль, чтобы учесть возможный отсев собак, не достигших критериев обучения. Собаки, не справившиеся с обучением задаче, были исключены (см. критерии в разделе ниже). В итоге пять самцов и пять самок в возрасте от 2 до 7 лет завершили полный эксперимент. Все собаки содержались в вольерах в Университетской ветеринарной клинике в Уппсале, Швеция, группами по 3–6 особей. Они содержались во внутренних вольерах площадью 24,3 м2 (9 м × 2,7 м) с 16:00 до 8:00 и во внешних вольерах площадью от 145 до 200 м2 (около 6 м × 25 м) в остальное время. Их кормили дважды в день кормом Hill’s Science Plan Mature Adult Chicken в количестве от 4 до 8 дл в день в зависимости от веса собаки, вода была доступна ad libitum. Дважды в неделю их выгуливал смотритель.
2. Стимулы для теста на предпочтение глобальных/локальных элементов
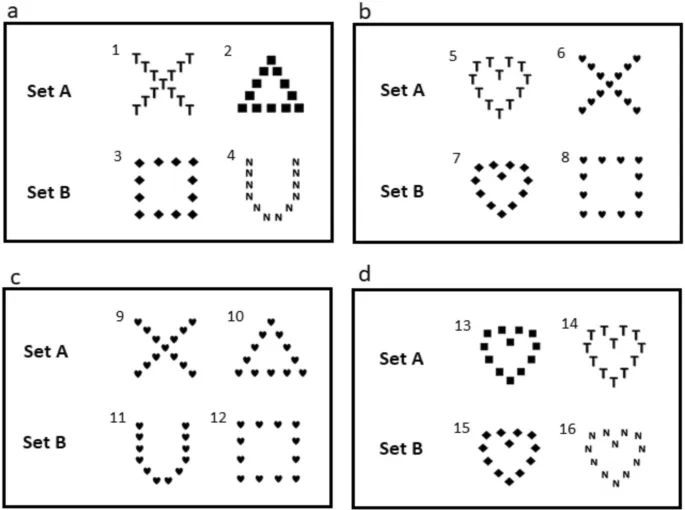
Тест на предпочтение глобальных/локальных элементов, использованный для оценки фокуса внимания, включал обучение собак различать различные визуальные стимулы, характеризующиеся двухуровневой иерархической структурой. Стимул состоял из глобальной формы, образованной меньшими локальными формами, что позволяло два уровня дискриминации: локальный или глобальный. Использованные нами стимулы были аналогичны стимулам Наво40. Локальные формы имели высоту и ширину 4 см, глобальные формы – 22,9 см. Стимулы были созданы в PowerPoint (Windows 10) с использованием черных непрозрачных символов на белом фоне. Были созданы два разных набора стимулов, чтобы избежать предвзятости, связанной с каким-либо конкретным аспектом паттерна стимулов41. Тренировочные стимулы показаны на Рис. 2а, и половина собак была случайным образом распределена в набор A, а другая половина – в набор B. Мы использовали функцию генератора случайных чисел в Microsoft Excel (функция = Rand()) для распределения собак по различным наборам символов. Конфликтные стимулы показаны на Рис. 2b, и они использовались для оценки локальных и глобальных предпочтений.
3. Экспериментальная установка
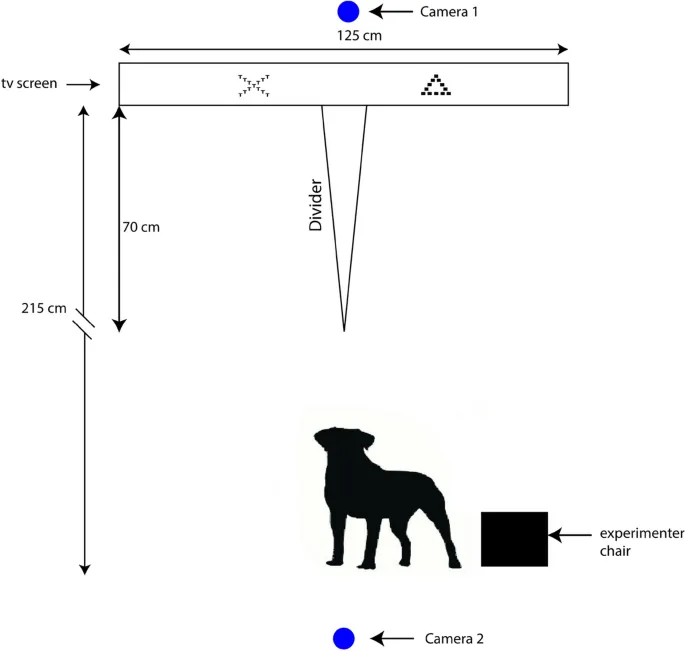
Эксперимент проводился в отдельной комнате, примыкающей к вольерам собак (1 минута ходьбы), и изначально была незнакома собакам. Собаки постепенно привыкали к комнате до начала обучения и тестирования. Тестовый аппарат состоял из экрана телевизора NEC Edition (длиной 125 см и высотой 72 см), нижний край экрана находился на высоте 10 см над уровнем пола. Динамик (Extron) был расположен непосредственно над экраном (на уровне середины экрана). Перед экраном телевизора была установлена перегородка длиной 70 см, разделяющая его на две половины, чтобы собаке было легче перейти на одну сторону или другую и коснуться носом только одного символа. Стул был расположен на расстоянии 2,15 м от экрана для исследователя, чтобы он мог сидеть во время обучения и тестирования собак (Рис. 3). Визуальный угол (визуальный угол = 2 * arctan (0,5 * размер объекта / расстояние)) от стартовой позиции составлял 5,58°, а визуальный угол от начала перегородки (70 см перед экраном) – 28,6°, при размере объекта 22,9 см).
4. Обучение
4.1. Привыкание
Собак сначала приучали к исследователям в их домашних вольерах, а затем к тестовой комнате. Затем их научили ассоциировать щелчок стандартного кликера с пищевым вознаграждением (смесь вареного ветчины и сухих собачьих бисквитов в соотношении 1 к 3). После этого собак обучали касаться носом одного вознаграждаемого стимула (см. Рис. 1а), напечатанного на бумаге формата А4, сначала на полу, а затем на стене, используя кликер-тренинг. Собак не лишали пищи перед обучением или тестированием.
4.2. Обучение тесту на предпочтение глобальных/локальных элементов
Собак обучали либо набором A, либо набором B в сессиях по 20 проб, максимум три раза в день, пять дней в неделю. В каждой сессии вознаграждаемый символ отображался 10 раз слева и 10 раз справа экрана в псевдослучайном порядке (не более трех раз подряд с одной стороны). Собаки должны были дать 16 из 20 правильных ответов в течение двух последовательных дней, чтобы перейти к следующему этапу обучения.
Обучение тесту на предпочтение глобальных/локальных элементов было разделено на четыре фазы, чтобы постепенно научить собак различать различные символы. В фазе 1 вознаграждаемый символ отображался с одной стороны экрана телевизора, а другая сторона оставалась пустой. В фазе 2 вознаграждаемый символ отображался с одной стороны экрана, а с другой стороны одновременно отображался один локальный элемент невознаграждаемого символа. В фазе 3 вознаграждаемый символ отображался против невознаграждаемого символа, который был уменьшен до 50% от своего полного размера. В тренировочной фазе 4 оба вознаграждаемых и невознаграждаемых символа отображались в полном размере (Рис. 1а). Максимальное количество сессий в каждой тренировочной фазе не было установлено, но мы останавливали обучение собак, не достигших критериев обучения, когда десять собак, успешно достигших критериев, завершили полный эксперимент.
В начале каждого испытания собаку держал за ошейник экспериментатор, сидящий на стуле. Каждое испытание начиналось с отображения знака плюс (шириной 1,7 см и высотой 2,1 см) в центре экрана в сочетании со звуком «плин» для привлечения внимания собаки к экрану. Если собака не смотрела на экран в течение примерно 10 секунд, звук повторялся один раз. Как только собака смотрела на экран, экспериментатор закрывал глаза, а затем одновременно отображались вознаграждаемый и невознаграждаемый символы (Рис. 1а). Примерно через 2 секунды экспериментатор говорил «иди» и отпускал собаку. Экспериментатор держал глаза закрытыми до тех пор, пока собака не отпускалась, чтобы избежать влияния на последующий выбор собаки. Экспериментатор снова открывал глаза, как только собака начинала двигаться к экрану. Собака получала подтверждающий звук кликера, а затем пищевое вознаграждение от экспериментатора, если подходила к экрану и касалась вознаграждаемого символа. Если собака подходила и касалась невознаграждаемого символа, щелчка и вознаграждения не давали, и испытание немедленно начиналось заново. Эта же процедура затем повторялась для других испытаний.
5. Тест на предпочтение глобальных/локальных элементов после различных воздействий
Все тестирование проводилось между 9:00 и 11:00. Перед началом теста каждая собака должна была правильно ответить на 4 из 5 финальных тренировочных проб (80% правильных ответов). Если собака не проходила этот критерий, она получала еще 5 тренировочных проб для прохождения критерия. Если тренировочный критерий был пройден, собака подвергалась воздействию, призванному индуцировать определенное эмоциональное состояние. Каждый эксперимент был разделен на два повторения: собаки сначала проходили тестирование во всех четырех эмоциональных воздействиях в случайном порядке (с использованием генератора случайных чисел Excel) в повторении 1, а затем это повторялось снова с использованием того же случайного порядка в повторении 2. Все собаки завершили повторение 1 перед тестированием в повторении 2. Причина повторения заключалась в балансировке стороны вознаграждаемого символа на экране, чтобы избежать смешения результатов из-за предвзятости к стороне или предпочтения одной стороны экрана/комнаты, если таковые могли существовать. Таким образом, при каждом эмоциональном воздействии вознаграждаемый символ отображался один раз слева и один раз справа экрана в случайном порядке в конфликте, локальных и глобальных тестах на различие (см. ниже).
Собаки проходили не более одного теста в день. Экспериментаторы не могли быть слепыми к воздействиям, поскольку они должны были быть применены непосредственно перед тестированием.
5.1. Эмоциональные воздействия
Четыре эмоции, которые мы стремились индуцировать, варьировались по валентности (позитивная, негативная) и уровню возбуждения (высокий, низкий) в двух различных контекстах вознаграждения, в соответствии с четырьмя квадрантами модели эмоций животных13, что привело к конкретным прогнозам в отношении фокуса внимания (см. Рис. 1). Позитивные состояния с высоким уровнем возбуждения создавались путем предоставления собаке небольшого количества желаемого30, но без возможности полного насыщения, чтобы вызвать (большее) желание или возбуждение (позитивное и высокое возбуждение). В Эксперименте 1 для индукции эмоций использовались пищевые вознаграждения. Мы использовали скрытую игрушку-головоломку («Dog Smart», Nina Ottosson, Швеция), состоящую из 9 отверстий с лакомствами, закрытых крышками. Собаки должны были убрать крышки ртом или лапами, чтобы получить доступ к лакомствам (Таблица 1).
Позитивные состояния с низким уровнем возбуждения индуцировались путем предоставления собакам вознаграждений до насыщения (социальный контекст) или до почти полного насыщения (пищевой контекст), чтобы создать состояние удовлетворения или contentment после потребления42. Не было возможности давать пищевые лакомства до тех пор, пока собаки не переставали их потреблять, потому что тогда они больше не были бы мотивированы выполнять саму задачу (что было выявлено как потенциальная проблема в пилотном исследовании до эксперимента).
Все позитивные воздействия были инициированы собакой, и собаки никогда не принуждались есть лакомства, играть или быть глаженными. Мы внимательно наблюдали за каждой собакой, чтобы определить, когда прекратить сессию в условиях низкого возбуждения в Эксперименте 2. Момент, когда собака вставала и уходила (что мы предполагали как потерю интереса) или когда собака полностью расслаблялась (некоторые собаки даже засыпали), служили точками для завершения воздействия и тестирования собаки. Этот временной момент варьировался от 15 до 40 минут, как указано в Таблице 2.
Негативные состояния с высоким уровнем возбуждения создавались путем ограничения доступа собак к желаемому вознаграждению, чтобы вызвать немедленную фрустрацию35. Негативные состояния с низким уровнем возбуждения индуцировались путем ожидания собакой вознаграждения, но никогда не предъявления этого вознаграждения (Таблицы 1 и 2). В условиях негативных состояний с низким уровнем возбуждения мы наблюдали за поведением каждой отдельной собаки и ждали, пока собаки перестанут искать вознаграждение. Затем мы тестировали собак через 30 секунд после потери интереса. Мы предположили, что эти воздействия сначала вызовут фрустрацию, но затем последует разочарование, когда они перестанут искать ожидаемое вознаграждение43, причем разочарование будет ниже по уровню возбуждения, чем фрустрация44.
Все собаки сначала завершили Эксперимент 1, прежде чем перейти к Эксперименту 2. Каждый эксперимент занимал около 8 дней (1 тест в день), то есть всего 16 дней для завершения полного исследования.
5.2. Конфликтный тест
Непосредственно после каждого эмоционального воздействия оценивались предпочтения собак в отношении локальной или глобальной информации в конфликтном тесте, примерно через 10–20 секунд после завершения воздействия. Это включало предъявление двух несовместимых тестовых стимулов (Рис. 2b): один символ с тем же глобальным элементом, что и у вознаграждаемого символа, но с локальным элементом, который никогда раньше не встречался (предпочтение которого указывало бы на глобальное предпочтение), и другой символ, показывающий те же локальные элементы, что и вознаграждаемый символ, но с глобальным элементом, который никогда раньше не встречался (предпочтение которого указывало бы на локальное предпочтение). Собаки всегда получали вознаграждение независимо от своего выбора. Каждая собака тестировалась дважды в каждом эмоциональном состоянии, а предъявление справа и слева глобальных и локальных выборов было сбалансировано по повторениям.
5.3. Тест на локальную и глобальную дискриминацию
Для подтверждения того, что собаки могли различать на локальном и глобальном уровнях, и обеспечения того, чтобы эмоциональные воздействия не ухудшали способность собаки различать на локальном и глобальном уровнях, непосредственно после конфликтного теста проводился один тест на локальную дискриминацию и один тест на глобальную дискриминацию20, с межурочным интервалом примерно 10–20 секунд. Процедура была идентична процедуре конфликтного теста, за исключением того, что правильный символ отличался от вознаграждаемого символа только на глобальном уровне для теста на глобальную дискриминацию (Рис. 2c) и только на локальном уровне для теста на локальную дискриминацию (Рис. 2d). Предъявление правильных локальных и глобальных стимулов справа и слева было рандомизировано, как и порядок локальной и глобальной дискриминации между двумя повторениями. Вознаграждался только выбор правильных локальных и глобальных символов.
5.4. Тесты мотивации
Тест на пищевую мотивацию и тест на социальную мотивацию проводились после завершения тестов на предпочтение глобальных/локальных элементов, чтобы оценить общий уровень пищевой и социальной мотивации каждой собаки. Это было сделано для учета индивидуальных различий в мотивации к пищевым и социальным стимулам45,46, которые потенциально могли повлиять на то, как собаки реагировали на эмоциональные воздействия в Эксперименте 1 и 2 соответственно. Тестирование мотивации проводилось в беговой дорожке длиной 20 м. Для теста на пищевую мотивацию на одном конце беговой дорожки помещалась миска с едой, содержащая 1 кусок колбасы, 1 кусок ветчины и 2 куска сухого корма. Для теста на социальную мотивацию на конце беговой дорожки стоял знакомый человек и продолжал звать собаку по имени, пока собака не достигала его. Записывалось время, которое собака тратила на пробежку к миске или к человеку с противоположного конца беговой дорожки. Собаке разрешалось съесть лакомства или ее гладили знакомый человек в течение 20 секунд. Этот процесс повторялся четыре раза для каждого стимула. Порядок тестов пищевой и социальной мотивации был рандомизирован, но сбалансирован по всей группе.
5.5. Процедура регистрации сердечного ритма и вариабельности сердечного ритма
Сердечный ритм собак регистрировался с помощью часов Polar, модель V800 PolarSystems (© Polar Electro 2021), для получения неинвазивного измерения ЧСС и ВСР в реальном времени. Пояс с двумя электродами крепился на грудь собаки и подключался к записывающему устройству, похожему на часы. Для улучшения контакта кожи с электродами использовался электрогель. Для каждой собаки проводилась 15-минутная запись (базовая линия) перед тем, как собаки подвергались эмоциональным воздействиям и глобально-локальному тестированию. Базовые линии регистрировались в той же комнате, где проводилось обучение и тестирование, чтобы обеспечить запись базовых линий в той же среде, что и воздействия.
Собаки всегда были очень взволнованы при входе в комнату для обучения и тестирования, поэтому мы не могли записывать базовые линии непосредственно перед проведением эмоциональных воздействий и тестов. Следовательно, запись базовых линий начиналась примерно через 5 минут после окончания тренировочной сессии, чтобы обеспечить состояние покоя, во время которого экспериментатор спокойно сидел, чтобы собаки расслабились и отдохнули. ЧСС и ВСР затем также регистрировались во время каждого эмоционального воздействия в течение первого повторения, поскольку, в отличие от теста на фокус внимания, эти показатели не должны были подвергаться влиянию потенциального предпочтения стороны.
Анализ данных
6.1. Данные о предпочтении глобальных/локальных элементов
Анализируемой переменной был выбор, сделанный в тесте на предпочтение глобальных/локальных элементов. Следовательно, это была биномиальная переменная (0 = выбор глобального; 1 = выбор локального), и анализ проводился по доле локальных выборов. Обобщенная линейная смешанная модель (GLMM) с логистической связью была подобрана с использованием пакетов lme449 и nmle50 с валентностью, возбуждением, повторением и их взаимодействиями, стороной предъявления локального выбора (слева или справа), назначенной группой стимулов (набор A или B) и полом собаки в качестве фиксированных эффектов, а идентификатором собаки – в качестве случайного эффекта. Для учета потенциальных индивидуальных различий в мотивации к пищевым или социальным стимулам мы включили среднее количество секунд, затраченное на достижение конца беговой дорожки в тестах пищевой и социальной мотивации, в качестве ковариат в модель, в Эксперименте 1 и 2 соответственно.
Из-за большого количества потенциальных фиксированных эффектов, которые могли повлиять на долю локальных выборов (например, валентность, возбуждение, повторение, их взаимодействия, пол, мотивация, сторона и символ стимулов), мы использовали информационный критерий для выбора модели. Значения критерия Акаике (скорректированные для малых выборок, AICc) были рассчитаны с помощью функции «dredge» пакета Mumin. Фиксированные эффекты в топовых моделях (с дельтой 0) были включены в финальный статистический анализ. Топовая модель в Эксперименте 1 включала взаимодействие валентности и повторения, а также основной эффект пола (AICc = 98,1, дельта = 0, вес = 0,312). Топовая модель в Эксперименте 2 включала основные эффекты валентности и пола, а также мотивацию в качестве ковариаты (AICc = 110,3, дельта = 0, вес = 0,038). Пост-хок тесты с коррекцией Тьюки проводились с пакетом emmeans. Данные социального теста показали избыточную дисперсию, поэтому для учета этого был включен случайный эффект на уровне наблюдения51.
Для сравнения способности собак различать на локальном или глобальном уровне, была проведена обобщенная линейная смешанная модель (GLMM) с логистической связью для сравнения долей правильных (= 1) и неправильных (= 0) выборов для теста на локальную и глобальную дискриминацию, с тестом на дискриминацию (локальный против глобального) в качестве фиксированного эффекта и ID собаки в качестве случайного эффекта.
6.2. Тест мотивации
Остатки от теста мотивации не имели нормального распределения, поэтому мы оценили, существует ли разница в скорости бега между социальными и пищевыми стимулами, а также между полами с помощью непараметрического теста Уилкоксона.
6.3. Сердечный ритм (ЧСС) и вариабельность сердечного ритма (ВСР)
Исходные данные были разделены на два временных периода для каждого воздействия (одна точка данных на собаку и период): 15-минутный базовый период и 45-секундный период воздействия в первом повторении (последние 45 секунд воздействия). Анализ ВСР требовал идентификации выбросов. Записи сначала проверялись алгоритмом AVEC (Algorithmsupported visual error correction) в R studio для выделения выбросов, которые затем вручную удалялись в Excel 2016 (Windows 10). В среднем удалялось 0,64% записей.
Очищенные записи сердечного ритма затем анализировались с помощью ARTiiFACT (программное обеспечение для обработки артефактов сердечного ритма и анализа вариабельности сердечного ритма, версия 2.1352), которое присваивало список индексов ВСР каждому временному периоду каждого воздействия. Анализ программного обеспечения ARTiiFACT дает все общие показатели ВСР, управляемой блуждающим нервом, во временном и частотном доменах (с использованием метода FFT), включая стандартное отклонение нормальных интервалов RR (SDNN), среднеквадратичное отклонение последовательных разностей (RMSSD), средние интервалы RR, долю последовательных интервалов R-R, которые отличаются более чем на 50 мс (pNN50), абсолютную мощность в высокочастотной полосе (HF), абсолютную мощность в низкочастотной полосе (LF) и абсолютную мощность в очень низкочастотной полосе (VLF). Диапазон VLF был установлен на 0,06 Гц, LF – на 0,24 Гц, а HF – на 1,04 Гц на основе измерений, проведенных у собак53.
Эти индексы анализировались в R с помощью GLMM, используя пакеты lme449 и lmerTest54, с эмоциональными воздействиями и полом в качестве фиксированных эффектов, базовыми значениями в качестве ковариаты и ID собаки в качестве случайного эффекта. Данные были логарифмически преобразованы, а предположения о нормальности и гомоскедастичности оценивались путем графического анализа остатков (пакет LMERConvienienceFunction55). Выбросы с остатками более 2,5 единиц были исключены из набора данных (содержащего 119 точек данных). В Эксперименте 1 мы исключили 1 выброс из SDNN, 1 выброс из LF и 1 выброс из VLF. В Эксперименте 2 мы исключили 1 выброс из VLF. Пост-хок тесты с коррекцией Тьюки проводились с использованием пакета emmeans47.
Результаты
Только 10 из 21 собаки прошли все критерии обучения. Эти 10 собак в среднем были полностью обучены после 364 ± 27 (среднее ± SE) проб, что эквивалентно 18 ± 1 (среднее ± SE) сессиям по 20 проб каждая.
В тесте мотивации средняя скорость бега по беговой дорожке была выше для социального вознаграждения (7,5 ± CI 3,9 с), чем для пищевого вознаграждения (12,3 ± CI 4,4 с, Wilcoxon V = 460,5, P = 0,005).
Эксперимент 1: Пищевые вознаграждения
7.1. Тест на предпочтение глобальных/локальных элементов
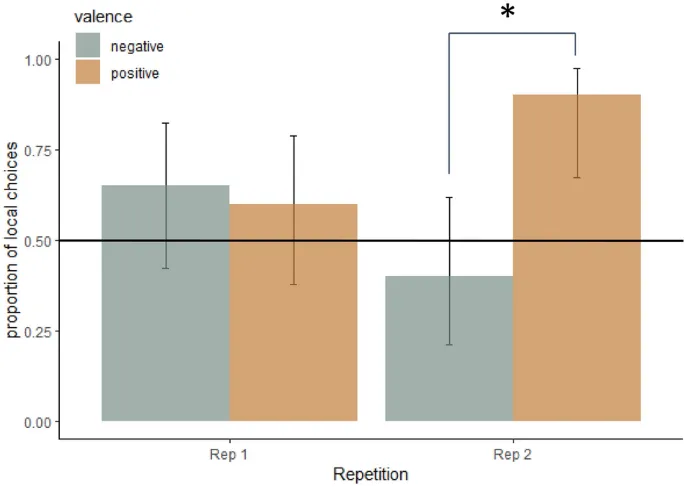
Наблюдалось значимое взаимодействие валентности и повторения по доле локальных выборов (F(1,7.2) = 7,2, P = 0,03, Рис. 4), при этом позитивные воздействия приводили к большему количеству локальных выборов по сравнению с негативными воздействиями во втором повторении (пост-хок тест Тьюки: P = 0,002).
Не было значимого взаимодействия валентности и возбуждения. Наблюдалась тенденция к влиянию пола на долю локальных выборов: самки собак чаще выбирали локальный символ (среднее 0,78, нижний CI = 0,62, верхний CI = 0,87) по сравнению с самцами (среднее 0,5, нижний CI = 0,35, верхний CI = 0,64, F(1,5) = 4,9, P = 0,08). Ни один из следующих переменных; возбуждение, набор тестовых стимулов (A или B), сторона предъявления (слева или справа) или скорость бега животного в тесте пищевой мотивации не повлиял на долю локальных выборов.
Мы также определили, отличался ли выбор при каждом воздействии от выбора 50–50 (указывающего на отсутствие предпочтения локальных или глобальных элементов), рассчитав доверительные интервалы (CI). Если CI не включают 0,5, то можно заключить, что имело место предпочтение локальной или глобальной информации. Мы обнаружили, что CI не включали 0,5 в условиях позитивной валентности во втором повторении (Рис. 4), предполагая, что позитивные условия действительно индуцировали предпочтение локальных элементов. Также, выборы самок не включали 0,5 в доверительном интервале (см. средние значения, приведенные ранее в этом разделе).
В тестах на дискриминацию, для оценки точности дискриминации на локальном и глобальном уровнях, собаки работали выше шансов как на локальном уровне (доля правильных выборов 0,78, с нижним CI 0,64 и верхним CI 0,87), так и на глобальном уровне (доля правильных выборов 0,65, с нижним CI 0,50 и верхним CI 0,77). Хотя численно собаки показали лучшие результаты в тесте на локальную дискриминацию, эта разница не была статистически значимой.
7.2. Сердечный ритм и вариабельность сердечного ритма
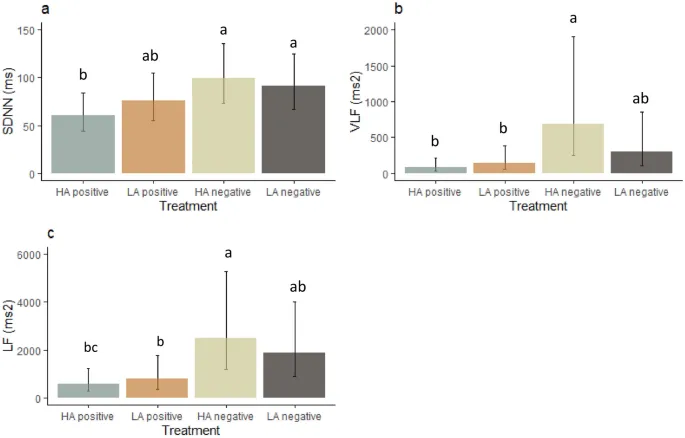
ВСР регистрировалась только в первом повторении. На SDNN влияло эмоциональное воздействие (F(3, 25,3) = 4,8, P = 0,009, Рис. 5a). Пост-хок анализ показал, что SDNN был ниже в условиях позитивного состояния с высоким уровнем возбуждения, чем в условиях негативного состояния с высоким уровнем возбуждения (P = 0,009) и негативного состояния с низким уровнем возбуждения (P = 0,036).
VLF также находился под влиянием эмоционального воздействия (F(3, 25,1) = 5,5, P = 0,005, Рис. 5b). В условиях негативного состояния с высоким уровнем возбуждения VLF был выше, чем в условиях позитивного состояния с высоким уровнем возбуждения (P = 0,0041) и позитивного состояния с низким уровнем возбуждения (P = 0,04). Кроме того, VLF имел тенденцию быть выше у самок (4476 ± 466 мс2) по сравнению с самцами (2796 ± 305 мс2, F(1, 6,7) = 4,1, P = 0,08).
LF зависел от воздействия (F(3, 25,4) = 6,6, P = 0,002, Рис. 5c). Пост-хок анализ выявил, что LF был выше в условиях негативного состояния с высоким уровнем возбуждения, чем в условиях позитивного состояния с высоким уровнем возбуждения (P = 0,004) и позитивного состояния с низким уровнем возбуждения (P = 0,03). Кроме того, в условиях негативного состояния с низким уровнем возбуждения LF также был выше, чем в условиях позитивного состояния с высоким уровнем возбуждения (P = 0,02). Никакие другие меры ВСР не находились под влиянием воздействий.
Эксперимент 2: Социальные стимулы
8.1. Тест на предпочтение глобальных/локальных элементов
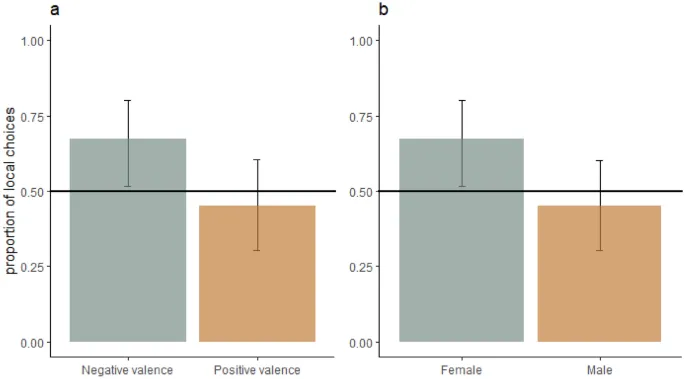
Все собаки играли с экспериментатором в условиях позитивного состояния с высоким уровнем возбуждения и хотели быть поглаженными в условиях позитивного состояния с низким уровнем возбуждения. Ни один из факторов, включенных в смешанную модель, статистически значимо не повлиял на долю локальных выборов (т. е. валентность, возбуждение, повторение, пол, сила социальной мотивации, набор и сторона, Рис. 6). Однако CI не включали долю локального выбора 0,5 в условиях негативной валентности, предполагая, что эти условия индуцировали предпочтение локальных элементов (Рис. 6). Также, CI у самок не включали долю локального выбора 0,5, предполагая, что самки чаще делали локальный выбор. Данные для каждого эмоционального воздействия можно увидеть на Рис. 2 в дополнительной информации.
При оценке дискриминации на локальном и глобальном уровнях только, собаки показали значительно лучшие результаты в локальной дискриминации (доля правильных ответов 0,84, с нижним CI 0,74 и верхним CI 0,90 правильных ответов) по сравнению с тестом на глобальную дискриминацию (0,63 правильных ответов, с нижним CI 0,51 и верхним CI 0,72 правильных ответов (F(1, 8,8) = 8,8, P = 0,01).
8.2. Сердечный ритм и вариабельность сердечного ритма
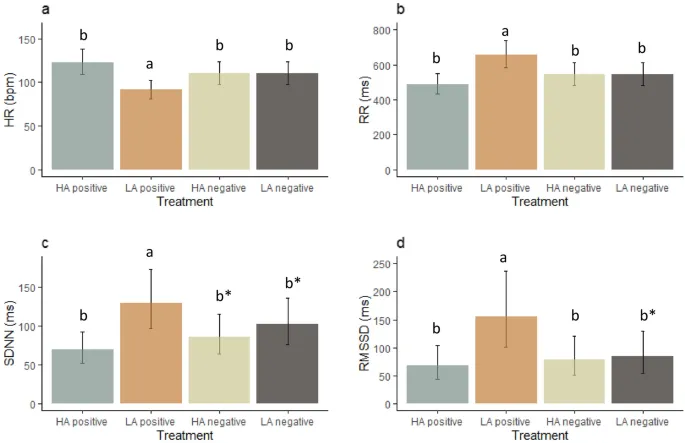
ЧСС находился под влиянием воздействия (F(3, 27) = 12,7, P < 0,001, Рис. 7a). Пост-хок анализ выявил, что ЧСС собак был ниже в условиях позитивного состояния с низким уровнем возбуждения, чем в условиях позитивного состояния с высоким уровнем возбуждения (P < 0,001), негативного состояния с высоким уровнем возбуждения (P = 0,037) и негативного состояния с низким уровнем возбуждения (P = 0,0039).
RR также находился под влиянием воздействия (F(3, 27) = 12,7, P < 0,001, Рис. 7b). Пост-хок тесты показали, что реакция RR собаки была выше в условиях позитивного состояния с низким уровнем возбуждения по сравнению с условиями позитивного состояния с высоким уровнем возбуждения (P < 0,001), негативного состояния с высоким уровнем возбуждения (P = 0,037) и негативного состояния с низким уровнем возбуждения (P = 0,039).
SDNN также находился под влиянием воздействия (F(3, 27) = 6,5, P = 0,003, Рис. 7c). У собак SDNN был выше в условиях позитивного состояния с низким уровнем возбуждения, чем в условиях позитивного состояния с высоким уровнем возбуждения (P = 0,002), и имел тенденцию быть выше, чем в условиях негативного состояния с высоким уровнем возбуждения (P = 0,06) и негативного состояния с низким уровнем возбуждения (P = 0,07).
RMSSD находился под влиянием воздействия (F(3, 27) = 5,0, P = 0,007, Рис. 7d). RMSSD был выше в условиях позитивного состояния с низким уровнем возбуждения, чем в условиях позитивного состояния с высоким уровнем возбуждения (P = 0,007) и негативного состояния с высоким уровнем возбуждения (P = 0,03), и имел тенденцию быть выше, чем в условиях негативного состояния с низким уровнем возбуждения (P = 0,06).
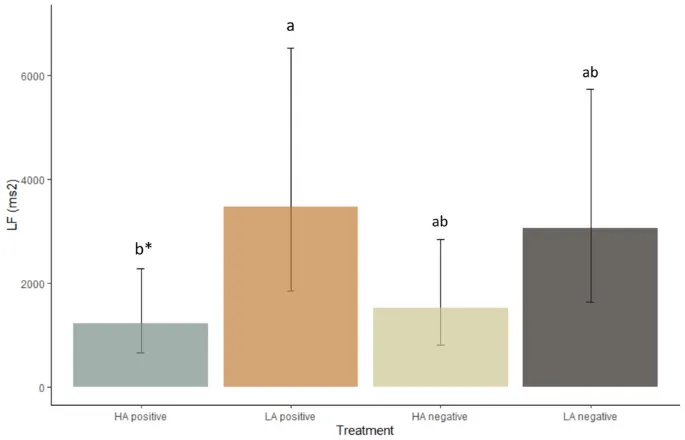
LF также находился под влиянием воздействия (F(3, 27) = 3,1, P = 0,04, Рис. 8). LF имел тенденцию быть выше в условиях позитивного состояния с низким уровнем возбуждения по сравнению с условиями позитивного состояния с высоким уровнем возбуждения (P = 0,07).
Обсуждение
Данное исследование нацелено на изучение фокуса внимания как нового индикатора эмоционального состояния у животных. В Эксперименте 1 мы обнаружили, что у собак в условиях позитивных воздействий, независимо от уровня возбуждения, наблюдалось сужение фокуса внимания по сравнению с собаками в условиях негативных воздействий, хотя это проявилось только во втором повторении. В Эксперименте 2 мы не выявили значимого влияния воздействий на сужение внимания, хотя собаки демонстрировали некоторую степень сужения внимания в негативных условиях по сравнению с позитивными. Хотя результаты не полностью соответствуют нашим ожиданиям, это первое свидетельство того, что эмоциональное состояние может изменять локальные и глобальные предпочтения у животных, как было показано ранее у людей. Возможно, индуцированные уровни возбуждения в некоторых условиях были не такими, как предполагалось (например, были высокими, а не низкими), и это может объяснить, почему результаты не всегда соответствовали прогнозам. Показатели ВСР подтвердили, что позитивные воздействия в Эксперименте 1 индуцировали предполагаемые позитивные эмоциональные состояния, хотя в Эксперименте 2 они могли отражать разницу в физической активности.
9.1. Фокус внимания
9.1.1. Эксперимент 1. Тест на пищевое вознаграждение
Во втором повторении внимание собак сужалось в условиях позитивной валентности, независимо от того, было ли это начальным или продолжительным доступом к пищевым вознаграждениям. Следовательно, представляется, что изменение фокуса внимания в основном отражало разницу в эмоциональной валентности, независимо от уровня возбуждения. Сужение внимания при ожидании пищевых лакомств (позитивное состояние с высоким уровнем возбуждения) подтверждается данными, полученными у людей4,9,56. Однако мы ожидали, что фокус внимания расширится при продолжительном доступе к пище. Одним из возможных объяснений отсутствия разницы между двумя условиями позитивной валентности может быть то, что собаки поддерживали высоко возбужденное состояние ожидания даже после получения множества пищевых лакомств, и нам, возможно, не удалось индуцировать позитивное состояние с низким уровнем возбуждения. Это подтверждается отсутствием разницы в ЧСС между позитивными условиями с низким и высоким уровнем возбуждения. Однако наша выборка была относительно небольшой из-за высокого процента отсева в исследовании, и поэтому мы должны интерпретировать наши результаты с осторожностью.
Негативное воздействие с высоким уровнем возбуждения (фрустрированный подход) не привело к прогнозируемому сужению внимания по сравнению с негативным воздействием с низким уровнем возбуждения, и собаки не показали явного предпочтения локальной или глобальной информации в негативных условиях. Возможно, негативное состояние с низким уровнем возбуждения не оказало достаточно сильного влияния на эмоциональное состояние собак, и поэтому мы не наблюдали четкого локального или глобального предпочтения. Кроме того, мы, возможно, не смогли снизить возбуждение в этом состоянии, и собаки могли все еще испытывать фрустрацию из-за отсутствия вознаграждения43,57. Несмотря на то, что параметры ВСР были численно ниже в условиях низкого возбуждения по сравнению с условиями высокого возбуждения, эти различия не достигли статистической значимости. Другое потенциальное объяснение заключается в том, что сам тест мог быть вознаграждающим для собак. Собаки были заметно взволнованы при входе в тестовую комнату и были очень мотивированы выполнять обучение и тесты. Поэтому эффект любого негативного воздействия мог исчезнуть, как только собаки выполнили конфликтный тест, который всегда вознаграждался.
В отличие от второго повторения, у собак не было четкого предпочтения локальных или глобальных элементов в любом эмоциональном состоянии в первом повторении. Эта временная разница могла быть связана с тем, что конфликтные стимулы ранее не предъявлялись собакам, и собаки могли быть сбиты с толку незнакомыми символами, т. е. реакция на новизну, маскирующая первоначальное предпочтение58. Эта возможность подкрепляется отсутствием подобной разницы между повторениями в Эксперименте 2, когда все собаки ранее сталкивались с конфликтными стимулами. В будущих экспериментах поэтому может быть целесообразно познакомить собак с конфликтными стимулами в рамках их обучения, чтобы уменьшить любое потенциальное влияние новизны.
9.1.2. Эксперимент 2. Тест на социальное взаимодействие
В Эксперименте 2 не было значимого влияния воздействия на сужение внимания. Доля локальных выборов в негативных условиях была выше 50% (т. е. CI не перекрывались с 0,5), предполагая некоторую степень сужения внимания. Учитывая сужение внимания после позитивных эмоций, которое мы наблюдали в Эксперименте 1, это кажется весьма противоречивым. Одним из возможных объяснений может быть то, что мы непреднамеренно создали негативное воздействие с высоким, а не низким уровнем возбуждения. Например, возможно, что отсутствие игры вызвало фрустрацию — поскольку собаки могли ожидать игры, но затем были проигнорированы — что привело к некоторой степени сужения.
Интересно, что мы не наблюдали подобного сужения внимания в позитивных условиях после социальных стимулов. Все собаки играли с экспериментатором, хотя, возможно, можно утверждать, что для собак более вознаграждающим является игра с человеком, к которому они социально привязаны (например, владелец или другая собака)59, а не со знакомым экспериментатором. В беговой дорожке собаки бежали быстрее к экспериментатору, чем к миске с едой. Это предполагает, что экспериментатор воспринимался как вознаграждающий, что подтверждается другими исследованиями33,60. Однако более быстрая скорость бега к экспериментатору, возможно, связана с тем, что он продолжал звать собаку, пока та не достигала его, в то время как миска с едой ставилась только один раз без дополнительного поощрения. Социальная мотивация поэтому не может быть напрямую сопоставлена с мотивацией к пище в нашем исследовании. Кроме того, стимулы, использованные для создания высокого (игра) и низкого (поглаживание) позитивного воздействия в социальном тесте, не были точно такими же (как в Эксперименте 1), хотя они и были связаны. Это также может быть причиной различий в результатах между Экспериментом 1 и 2.
В обоих экспериментах собаки показали лучшие результаты в тесте на дискриминацию на локальном уровне, чем на глобальном, хотя в Эксперименте 1 это не достигло статистической значимости. Поэтому собаки в нашем исследовании, казалось, имели общее предпочтение локальных элементов и лучшую способность к дискриминации на локальном уровне. Предпочтение локальной информации над глобальной было обнаружено у ряда видов21,22,23, хотя ранее у собак не было выявлено четкого предпочтения19. Локальное предпочтение в нашем исследовании может быть связано с небольшой выборкой, или с индивидуальными предпочтениями19, или, возможно, с конкретной породой, которую мы использовали. Бигль был изначально выведен для охоты. Селекционное разведение могло привести к высокой концентрации внимания на деталях окружающей среды, чтобы облегчить поимку добычи, что может объяснить общее предпочтение локальных стимулов. Также возможно, что пространственная плотность наших элементов была слишком низкой, что также могло усилить предпочтение локальных элементов22,61. Следовательно, в будущих исследованиях может быть возможно улучшить дискриминацию на глобальном уровне, увеличив плотность символов.
Самки собак имели тенденцию к более сильному предпочтению локальных элементов, чем самцы, в обоих экспериментах. Эти результаты подтверждаются другими исследованиями, в которых женщины демонстрировали лучшую способность обрабатывать локальные элементы62. Эти половые различия могут быть обусловлены асимметрией полушарий, причем правое полушарие обрабатывает глобальные элементы, а левое полушарие — локальные элементы более эффективно62,63. Однако в предыдущих исследованиях у собак не было выявлено эффекта пола на локальные/глобальные предпочтения19,64.
Задача обучения на глобальные/локальные элементы была сложной, и только 10 из 21 собаки прошли все критерии обучения. Некоторые собаки были слишком напуганы и не подходили к экрану. Другие научились подходить к стимулам, но не прошли критерии обучения. Мы предполагаем, что возраст собаки играл роль, поскольку ни одна собака старше 8 лет не достигла критериев. Хорошо известно, что когнитивные способности резко снижаются после 8 лет65, и это может быть причиной того, что мы не смогли обучить более старых собак этой сложной задаче. Кроме того, собаки, использованные в этом эксперименте, были лабораторными собаками, содержащимися в вольерах, и не были знакомы с обучением визуальной дискриминации, и обучение задаче было для них сложным. Однако высокий процент отсева во время обучения также сообщался в предыдущем исследовании на домашних собаках19. Мы рекомендуем использовать гораздо большую выборку в будущих исследованиях для учета высокого процента отсева.
9.2. Сердечный ритм и вариабельность сердечного ритма
Для получения дополнительных данных, которые могли бы подтвердить, индуцировали ли мы предполагаемые эмоциональные состояния, мы также регистрировали показатели ЧСС и ВСР в первом повторении каждого эксперимента. SDNN является хорошим предиктором общей ВСР во время записи28. SDNN был ниже в позитивном состоянии с высоким уровнем возбуждения по сравнению с обоими негативными состояниями, что предполагает, что получение лакомств действительно воспринималось собаками как позитивное. Состояние низкого возбуждения было несколько промежуточным. LF указывает на симпатическую активность, а VLF – на симпатовагальный баланс28,66, и эти показатели также коррелируют с SDNN67. Как LF, так и VLF были ниже в позитивных условиях, что предполагает активацию симпатической нервной активности и ингибирование вагальной активности66,68, связанное с позитивной валентностью. Это согласуется с предыдущим исследованием, в котором LF был ниже, когда собаки ожидали тефтель, по сравнению с менее желательным стимулом30. Это подтверждает, что наши позитивно окрашенные условия действительно воспринимались собаками как позитивные.
В Эксперименте 2 состояние низкого возбуждения с позитивной валентностью снижало ЧСС по сравнению с другими состояниями, в то время как оно увеличивало RR, SDNN, RMSS и LF. RMSSD отражает вагальную тональную регуляторную активность28 и может отражать валентность у собак30,33. Однако увеличение показателей ВСР в нашем исследовании, скорее всего, связано с отсутствием физической активности, связанной с поглаживанием, по сравнению с другими воздействиями69,70,71. Тем не менее, это также предполагает, что сеанс поглаживания мог восприниматься как успокаивающий и, следовательно, с более низким уровнем возбуждения по сравнению с другими воздействиями30,33,72. Следовательно, индексы ВСР, казалось, отражали изменение физической активности в нашем исследовании и, возможно, снижение возбуждения.
Временные периоды, в течение которых мы регистрировали ЧСС и ВСР, были относительно короткими, и обычно рекомендуются периоды не менее 5 минут, особенно для частотных показателей28. Однако собаку очень трудно удерживать в состоянии возбуждения или фрустрации дольше минуты, поскольку она теряет возбуждение или перестает пытаться получить вознаграждение. Другие исследования предполагают, что периоды длительностью всего 1 минута достаточны для измерения ЧСС, SDNN и RMSSD при условии тщательного удаления артефактов73, что мы и сделали. Предыдущие исследования использовали еще более короткий промежуток времени для регистрации частотных показателей ВСР у собак (20 секунд)30. Более того, было показано, что сверхкраткосрочный анализ ВСР достаточен для оценки психического стресса74, причем даже частотные показатели дают надежные результаты в течение 40–50 секунд, что аналогично нашему исследованию (45 секунд). Однако мы должны быть осторожны при интерпретации частотных показателей из-за коротких временных периодов и небольшого размера выборки в нашем исследовании.
Было сложно получить соответствующую базовую линию для ВСР. Мы не могли записать базовую линию покоя в домашнем вольере, поскольку собаки содержались группами, и другие собаки могли повлиять на записи. Мы также не могли записать базовую линию в нейтральном эмоциональном состоянии непосредственно перед тестированием, потому что собаки всегда были очень взволнованы при входе в комнату для обучения и тестирования. Поэтому мы решили записывать базовые линии примерно через 5 минут после тренировочной сессии, когда собаки были расслаблены и отдыхали, но в той же комнате, что и обучение и тестирование, чтобы контролировать среду. В будущих экспериментах может быть целесообразно оценить и сравнить различные базовые линии ЧСС (например, домашний вольер, после тренировки и перед тестированием) с ЧСС во время воздействия.
Мы решили не контролировать физическую активность собак во время воздействий, чтобы не влиять на реакции собак в локальных/глобальных тестах. Однако, в то время как уровни активности в позитивных условиях с низким и высоким уровнем возбуждения, а также в негативных условиях с высоким уровнем возбуждения были примерно одинаковыми (т. е. собаки топтались и ходили вокруг скрытой пищевой игрушки или топтались и ходили вокруг стойки, закрывающей ее), физическая активность была ниже в социальном условии с низким уровнем возбуждения, и, возможно, также в пищевом условии с низким уровнем возбуждения и негативной валентностью.
Заключение
Данное исследование предоставляет первое указание на то, что эмоциональное состояние может изменять фокус внимания у животных. Позитивные эмоции, возникающие в результате потребления пищевых вознаграждений, приводили к сужению фокуса внимания по сравнению с негативными эмоциями, связанными с потерей пищевого вознаграждения. Показатели ВСР подтвердили, что получение пищевых вознаграждений было позитивным опытом по сравнению с недоступными пищевыми вознаграждениями, которые, вероятно, воспринимались собаками как фрустрирующие. Однако позитивные социальные взаимодействия не привели к сужению внимания таким же образом, как пищевые вознаграждения. Таким образом, хотя методология оценки фокуса внимания показывает определенные перспективы в выявлении различий в валентности, необходимы дальнейшие исследования с большей выборкой. Тем не менее, поскольку это новая методология измерения эмоций у животных, этот эксперимент может быть использован для лучшего прогнозирования в будущих исследованиях.





