Интересное сегодня
Введение в психофизические различия между фазовыми и позиционными сигналами зрительного движения
Движение в зрительных образах может быть описано в терминах изменения пространственного положения особенностей изображения с течением времени или как изменение фаз компонентов пространственной частоты Фурье изображения с течением времени. Хотя теоретически эти описания должны быть математически эквивалентными и сигнализировать о движении в одном направлении, можно создать стимулы кажущегося движения, в которых эти два описания разделяются и сигнализируют о движении в разных направлениях.
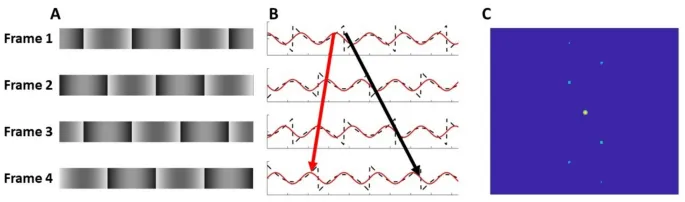
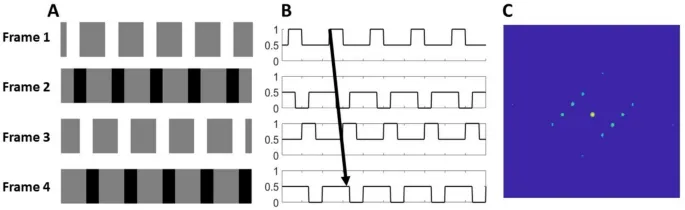
Например, решетка с отсутствующей фундаментальной гармоникой (рис. 1, меандровая решетка с удаленным компонентом основной пространственной частоты), смещенная на эквивалент одной четверти цикла отсутствующей фундаментальной гармоники, создает два конфликтующих восприятия: движение в направлении смещения, сигнализируемое изменением положения особенностей (т.е. границ с высоким контрастом или областей яркости), и движение в противоположном направлении, сигнализируемое наложением фазы следующей наиболее salient (третьей) гармоники. В зависимости от пространственных и временных параметров наблюдатели могут по умолчанию воспринимать последнее и "видеть" движение в направлении смещения только при целенаправленном внимании к особенностям и отслеживании их смещения.
Терминология и аналогии
Для краткости мы называем изменение фазы компонентов Фурье «фазовым сигналом», а изменение пространственного положения особенностей изображения — «позиционным сигналом». Фазовый сигнал аналогичен движению первого порядка, которое относится к движению, создаваемому дрейфующей модуляцией яркости, а позиционный сигнал аналогичен движению третьего порядка, которое относится к изменениям положения salient особенностей, как определено Лу и Сперлингом.
Мы используем термины «фазовый сигнал» и «позиционный сигнал», поскольку они описывают изменения в стимулах кажущегося движения между кадрами, без предположений о том, доступны ли эти сигналы системам, выполняющим анализ первого или третьего порядка. Обычно стимулы могут содержать множественные фазовые и позиционные сигналы, одновременно сигнализируя о движении во многих совпадающих и конфликтующих направлениях.
Нейронные корреляты фазовых и позиционных сигналов зрительного движения
Зрительная система полагается на два основных пути для передачи информации от сетчатки к зрительной коре. Парвоцеллюлярный путь берет начало от миджет ганглиозных клеток сетчатки, проецируется через парвоцеллюлярные пластинки дорсального латерального коленчатого ядра (dLGN) таламуса в слой 4Cβ первичной зрительной коры и составляет около 80% ганглиозных клеток сетчатки и релейных нейронов dLGN.
Магноцеллюлярный путь берет начало от парасольных ганглиозных клеток сетчатки, проецируется через магноцеллюлярные пластинки dLGN в слой 4Cα первичной зрительной коры и составляет около 10% ганглиозных клеток сетчатки и релейных нейронов dLGN.
Функциональные различия между путями
Функционально между этими двумя путями существуют важные различия. Нейроны парвоцеллюлярного пути имеют относительно небольшие рецептивные поля, избирательно реагируют на длину волны света и имеют устойчивые ответы, тогда как нейроны магноцеллюлярного пути имеют большие рецептивные поля, являются широкополосными (т.е. нечувствительными к длине волны света), имеют транзиентные ответы и более чувствительны к низкому контрасту.
Хотя два пути анатомически разделены вплоть до первичной зрительной коры и в некоторой степени через экстрастриарную кору, между ними существует определенное перекрытие в диапазоне пространственной и временной чувствительности, при этом система миджет клеток настроена на более высокие пространственные частоты и более низкие временные частоты, а система парасольных клеток предпочитает более низкие пространственные частоты и более высокие временные частоты.
Цели и гипотезы исследования
Мы рассматриваем вопрос, можно ли психофизически разделить чувствительность к фазовым и позиционным сигналам, и коррелирует ли производительность различения движения на основе разных сигналов с различиями в топографии магноцеллюлярных и парвоцеллюлярных субстратов.
Мы измерили пороги смещения для различения направления при четырех эксцентриситетах сетчатки в височном поле зрения (носальная сетчатка) между центральной ямкой и 15° у семи наблюдателей с использованием двухкадровых стимулов кажущегося движения Габора, представленных на семи пространственных частотах с процедурой временного выбора из двух альтернатив (2AFC).
Методология эксперимента
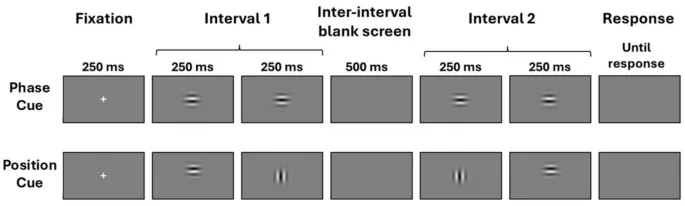
Фазовый сигнал генерировался путем смещения фазы синусоидальной несущей решетки на четверть цикла (90°) за статичным окном между кадрами, в то время как позиционный сигнал генерировался путем смещения гауссова окна на эквивалентное расстояние при устранении фазового сигнала путем поворота несущей на 90° между кадрами.
На каждом эксцентриситете порог смещения наблюдателя принимался за одну четверть цикла наивысшей пространственной частоты, при которой наблюдатель мог правильно сообщить о направлении движения в 75% испытаний. Затем мы сравнили наклоны этих психофизических порогов как функции эксцентриситета с опубликованными моделями топографии парасольных и миджет ганглиозных клеток в человеческой сетчатке, предположив, что они будут соответствовать порогам смещения для фазовых и позиционных сигналов соответственно.
Результаты: пороги различения направления движения
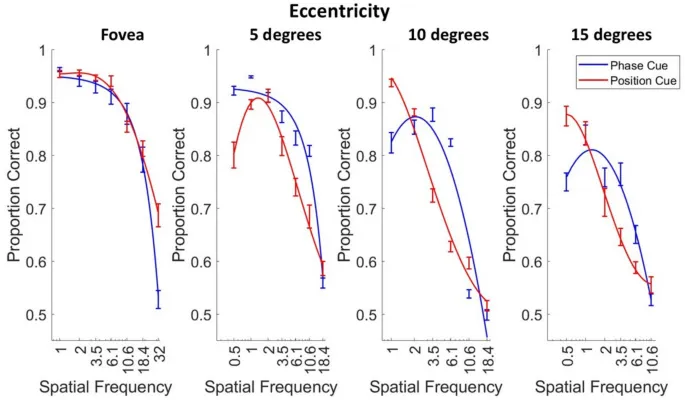
Сначала мы измерили способность семи наблюдателей различать направление движения, изображенное в двухкадровых кинематограммах, как функцию пространственной частоты, эксцентриситета сетчатки и того, было ли движение обозначено изменением фазы или положения.
Стимулы представляли собой патчи Габора, в которых фазовый сигнал генерировался путем смещения фазы синусоидальной несущей решетки на четверть цикла (90°) за статичным взвешенным по Гауссу окном, тогда как позиционный сигнал генерировался путем смещения взвешенного по Гауссу окна на эквивалентное расстояние при устранении фазового сигнала путем поворота несущей на 90° между кадрами.
Анализ высокочастотных срезов
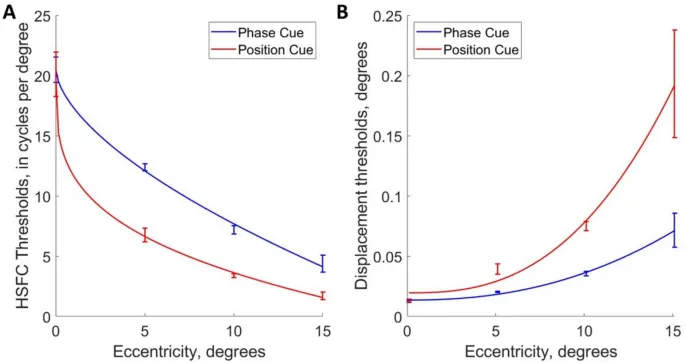
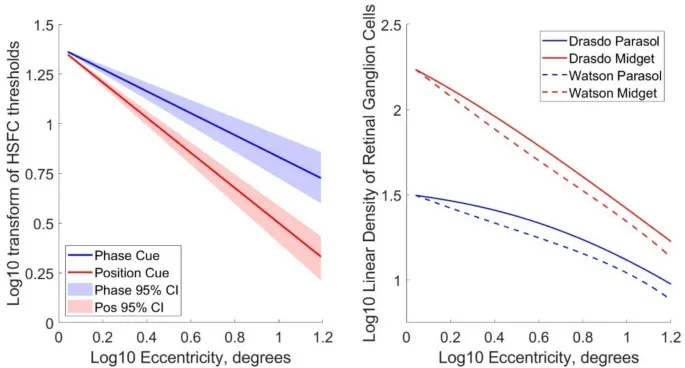
Затем мы интерполировали для каждого эксцентриситета для каждого наблюдателя наивысшую пространственную частоту, при которой наблюдатель правильно сообщал о направлении движения, изображенном в 75% испытаний. Эти данные суммированы на рис. 4A, который показывает, что эти высокочастотные срезы (HSFC) уменьшаются более круто с эксцентриситетом для различения направления по позиционному сигналу, чем для различения направления по фазовому сигналу.
Изменения высокочастотных срезов с эксцентриситетом аппроксимируются степенными функциями, которые при нанесении на логарифмические оси отображаются в виде прямых линий, где наклон соответствует показателю степенной функции.
Корреляция между высокочастотными срезами и плотностью ганглиозных клеток
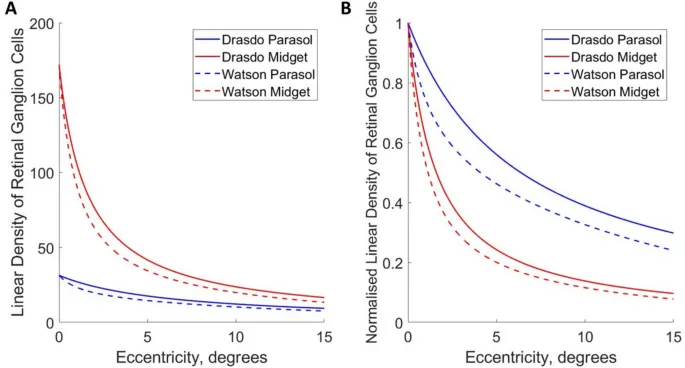
Модели плотности парасольных и миджет ганглиозных клеток между 0° и 15° эксцентриситета вдоль носовой сетчатки были получены из Watson (2014) и Drasdo (1989).
Как модели распределения плотности ганглиозных клеток, так и данные высокочастотных срезов наблюдателей могут быть аппроксимированы степенными функциями. Плотность ганглиозных клеток выражается как линейная плотность (ганглиозные клетки на градус, вычисленная как квадратный корень из планарной плотности), что уместно для стимулов пространственной частоты 1D, использованных в этом психофизическом эксперименте.
Сравнение наклонов регрессий
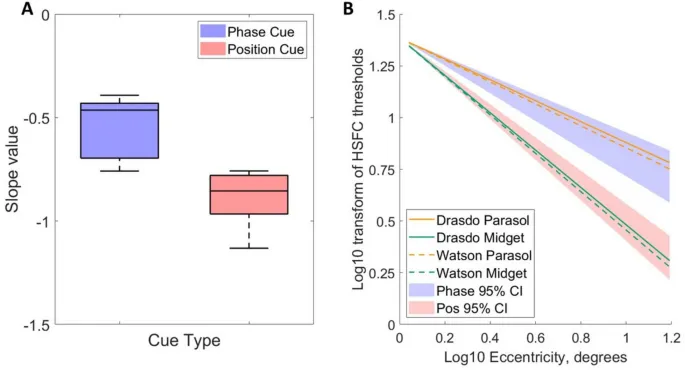
Наклоны линейных регрессий, подобранных к логарифмически преобразованным данным плотности ганглиозных клеток как функции эксцентриситета, сравнивались с наклонами высокочастотных срезов наблюдателей как функции эксцентриситета для фазового и позиционно обозначенного движения.
Наклоны моделей плотности парасольных ганглиозных клеток сетчатки, полученные из Watson (2014), наклон = -0.531, и из Drasdo (1989), наклон = -0.505, явно попадают в 95% доверительные интервалы наклонов высокочастотных срезов наблюдателей для фазового сигнала (средний наклон = -0.552, 95% ДИ [-0.696, -0.431]).
Наклоны моделей плотности миджет ганглиозных клеток (Watson (2014): наклон = -0.929; Drasdo (1989): наклон = -0.903) явно попадают в 95% доверительные интервалы наклонов высокочастотных срезов наблюдателей для позиционного сигнала (средний наклон = -0.884, 95% ДИ [-0.966, -0.779]).
Обсуждение результатов
Мы предположили, что более крутое снижение чувствительности к позиционным сигналам по сравнению с фазовыми сигналами с увеличением эксцентриситета сетчатки может свидетельствовать о том, что фазовые и позиционные сигналы обрабатываются (по крайней мере, изначально) разделяемыми нейронными субстратами, а именно парасольными и миджет ганглиозными клетками сетчатки (RGC).
В соответствии с этой гипотезой мы обнаружили, что высокочастотные срезы наблюдателей значительно круче снижались в условии позиционного сигнала, чем в условии фазового сигнала. Это повторяет предыдущие findings, показывающие периферическое смещение в сторону фазовых сигналов над позиционными сигналами в обработке движения, как показано в стимулах с конфликтующими сигналами.
Альтернативные объяснения и поддержка findings
Возможное альтернативное объяснение этих результатов — различный пространственный масштаб фазовых и позиционных сигналов. Хотя оба сигнала представлены в стимуле Габора, фазовый сигнал (смещение несущей) может восприниматься в масштабе одного цикла несущей, в то время как позиционный сигнал (смещение гауссова конверта) является "задачей центра тяжести", где из-за вращения несущей между кадрами изменение положения конверта требует интегрирования по всему стимулу.
Поддержка наших findings может быть найдена в предыдущих исследованиях, которые сравнивают градиенты порогов по сетчатке, которые, несмотря на использование разных стимулов, обнаружили similar gradients в снижении порогов пространственной частоты (или, если использовать пороги смещения, то увеличение порогов смещения с эксцентриситетом) к gradients, представленным в этой статье, при тестировании чувствительности к движению и положению.
Методология психофизического эксперимента
Семь наблюдателей (трое мужчин) в возрасте 21–57 лет с нормальным или скорректированным до нормального зрением были включены в это исследование. Все наблюдатели дали письменное информированное согласие перед участием в исследовании.
Аппаратура и стимулы
Стимулы представлялись на мониторе CRT (Mitsubishi Diamond Pro 2070, 1600 × 1200 пикселей) и гамма-корректировались с помощью колориметра ColorCAL MK II (Cambridge Research Systems) с максимальной яркостью 81.33 кд/м². Расстояние просмотра устанавливалось на 180 см от монитора для стимулов, рассматриваемых центрально и при эксцентриситете 5°, и на 110 см для стимулов, рассматриваемых при эксцентриситете 10° и 15°, и поддерживалось с помощью подставки для подбородка.
Двухкадровые кинематограммы кажущегося движения были построены из пар патчей Габора (синусоидальных решеток, взвешенных двумерным гауссовым окном) в MATLAB. Пространственная частота несущих решеток варьировалась от 0.3° до 32° со стандартным отклонением гауссова окна, установленным на 2.5 цикла длины волны.
Заключение и значение исследования
Мы предоставили доказательства того, что пороги различения направления движения на основе фазовых и позиционных сигналов значительно различаются в периферической сетчатке, причем пороги для движения, обозначенного положением, падают более круто, чем пороги для движения, обозначенного фазой.
Относительное изменение порогов с эксцентриситетом для фазовых и позиционных сигналов движения коррелирует с относительным изменением линейной плотности клеток парасольных и миджет ганглиозных клеток сетчатки соответственно. Это позволяет предположить, что магноцеллюлярный (парасольный) и парвоцеллюлярный (миджет) пути могут составлять разделяемые нейронные субстраты для движения первого порядка (Фурье) и движения третьего порядка (отслеживания особенностей).
Это исследование имеет важное значение для понимания фундаментальных механизмов зрительного восприятия и может найти применение в ранней диагностике патологических состояний, влияющих на различные классы ганглиозных клеток сетчатки.