
Интересное сегодня

Оптимизация когнитивного контроля: Влияние интенсивности и длительности стимуляции tDCS
Введение
Транскраниальная стимуляция постоянным током (tDCS) — это неинвазивная методика нейромодуляции, широко применяемая для исследования причинно-следственных связей между областями мозга и их когнитивными функциями. В последние десятилетия все больше исследований tDCS направлены на изучение нейронных коррелятов когнитивных функций путем изменения производительности в различных когнитивных задачах, таких как обработка языка (например, [1,2,3]), рабочая память (например, [4]) и приобретение моторных навыков (например, [5]). Однако лишь немногие исследования пролили свет на эффекты различных параметров tDCS, таких как интенсивность или длительность стимуляции [6,7,8].
Согласно предыдущим исследованиям [8,9], tDCS модулирует кортикальную возбудимость, регулируя потенциал покоя мембраны, что, в свою очередь, изменяет вероятность их активации. Анодная tDCS обычно усиливает кортикальную возбудимость, вызывая деполяризацию нейронов, что облегчает синаптическую передачу, в то время как катодная tDCS способствует гиперполяризации нейронов, снижая кортикальную возбудимость [10]. Согласно физике tDCS, интенсивность тока в мозге линейно увеличивается пропорционально приложенному току [11]. Более конкретно, эффекты tDCS зависят от различных параметров, включая интенсивность и длительность тока, которые, как теоретически предполагается, определяют степень изменений возбудимости [12]. Тем не менее, взаимосвязь между интенсивностью стимуляции и эффективностью стимуляции остается неопределенной. Например, Karuza et al. [13] обнаружили, что эффект помех в задаче Фланкера значительно снижался при стимуляции дорсолатеральной префронтальной коры (DLPFC) анодной tDCS с интенсивностью 1,5 мА. Напротив, Zmigrod и коллеги [14] исследовали влияние анодной tDCS с интенсивностью 2 мА на DLPFC на подавление помех с использованием задачи Фланкера, но не обнаружили значимых эффектов. На сегодняшний день лишь немногие исследования напрямую сравнивали эффекты различных интенсивностей стимуляции. В исследовании с использованием задачи принятия сенсомоторных решений Ehrhardt et al. [15] изучили влияние интенсивности тока (т.е. 0,7 мА, 1 мА и 2 мА) на принятие решений и обнаружили значительное улучшение как времени реакции, так и точности после tDCS с интенсивностью 1 мА, но не 0,7 или 2 мА. Другое исследование по моторному обучению предполагало значительное улучшение моторной производительности только после tDCS с интенсивностью 1,5 мА, но не 1 мА [16]. В целом, различные интенсивности стимуляции могут давать разные результаты, но полученные данные пока неубедительны.
Другим потенциальным фактором, влияющим на эффективность tDCS, является длительность стимуляции [9]. В исследовании с использованием задачи стоп-сигнала Mansouri и коллеги [17] изучили влияние анодной tDCS с интенсивностью 1,5 мА на DLPFC в течение 10 минут на когнитивный контроль и обнаружили значительное снижение времени реакции стоп-сигнала, что указывает на улучшение когнитивного контроля. Напротив, Stramaccia et al. [18] не наблюдали значительного улучшения при применении анодной tDCS с интенсивностью 1,5 мА к DLPFC в течение 20 минут перед той же задачей. Это расхождение в поведенческих результатах подчеркивает необходимость дальнейшего изучения влияния длительности стимуляции в рамках одного исследования.
Кроме того, взаимодействие между интенсивностью и длительностью tDCS не обязательно является кумулятивным. При более высоких интенсивностях длительная стимуляция может вызывать изменения в гомеостатической пластичности, которые потенциально могут обратить вспять намеченные возбуждающие эффекты. Таким образом, эффект стимуляции зависит не только от каждого параметра по отдельности, но и от того, как взаимодействие параметров влияет на кортикальную возбудимость с течением времени. Основываясь на приведенных выше физиологических и эмпирических данных, мы предположили, что исходы стимуляции будут различаться в зависимости от взаимодействия между интенсивностью и длительностью.
Предыдущие исследования дали противоречивые результаты относительно влияния интенсивности и длительности tDCS. Одной из возможных причин является то, что большинство предыдущих исследований полагались на традиционные меры, такие как время реакции (RT) и уровень точности, для оценки общей производительности. Влияние стимуляции на когнитивную производительность может приводить к улучшению различных подкомпонентов когнитивных процессов. Поэтому важно дополнительно разложить данные RT и точности. Модель диффузии дрейфа (DDM) — хорошо зарекомендовавшая себя вычислительная модель процессов принятия решений в задачах с вынужденным выбором из двух вариантов, — предоставляет количественную основу для объяснения человеческого поведения при принятии решений [19,20,21]. В рамках DDM процесс принятия решений теоретически разлагается на отдельные параметры, включая скорость дрейфа и время не принятия решения (для всестороннего обзора см. [22]). Эта попытка может быть полезной для выявления более тонких изменений при варьировании параметров tDCS, что, в свою очередь, способствует более точным параметрам tDCS как для клинических, так и для теоретических целей.
Кроме того, большинство исследований в основном применяли tDCS к одной области для установления причинно-следственных связей между областями мозга и когнитивными функциями (например, [1,23,24,25]). В отличие от доказанной эффективности односайтовой tDCS на моторной коре [26], получены противоречивые результаты относительно влияния односайтовой tDCS на более сложные когнитивные функции, такие как обработка языка (например, [27,28]) и когнитивный контроль (например, [3,29]). Учитывая, что сложные когнитивные функции зависят от взаимодействия и интеграции локальных и удаленных областей мозга через высокосвязанные сети [30], двухсайтовая анодная tDCS может увеличить кортикальную возбудимость больше, чем односайтовая [31]. Поэтому стоит исследовать потенциально различные эффекты односайтовой и двухсайтовой tDCS.
С этой целью в настоящем исследовании мы стремились изучить влияние различных интенсивностей и длительностей стимуляции анодной tDCS на когнитивную обработку, используя DDM наряду с традиционными поведенческими мерами. Кроме того, мы стремились исследовать различия в эффектах стимуляции, производимых односайтовой и двухсайтовой tDCS. Эта попытка может установить количественную зависимость между дозировкой tDCS и поведенческой производительностью. Задача Фланкера [32], часто используемая парадигма для оценки когнитивного контроля [33], была адаптирована. Было показано, что дорсолатеральная префронтальная кора (DLPFC) и дополнительная моторная кора (SMA) активируются сильнее при несогласованных пробах по сравнению с нейтральными пробами [34,35,36,37], а мозжечок также участвует в когнитивном контроле, связанном с задачей Фланкера [38,39,40,41]. Была продемонстрирована структурная связь между мозжечком и корой головного мозга [42]. Поэтому в настоящем исследовании были выбраны пре-дополнительная моторная кора (preSMA) (односайтовая стимуляция) и пре-дополнительная моторная кора – мозжечок (preSMA–мозжечок) (двухсайтовая стимуляция) в качестве целевых областей для исследования влияния tDCS на эффект помех в задаче Фланкера. В частности, в Эксперименте 1 мы изучили влияние tDCS с интенсивностью 1 мА и 1,5 мА на RT и параметры DDM, включая как скорость дрейфа, так и время не принятия решения [22,43]. Мы выбрали интенсивности 1,0 мА как низкую, а 1,5 мА как высокую, основываясь на предыдущей литературе, документирующей нейрофизиологические и когнитивные различия между этими двумя интенсивностями (например, [16,44,45,46]). Например, Khalil et al. (2023) исследовали влияние интенсивности tDCS на задачу восприятия, обозначив 1,0 мА как низкую интенсивность и 1,5 мА как высокую, и обнаружили, что производительность в задаче улучшилась только после tDCS с интенсивностью 1 мА, но не 1,5 мА. В Эксперименте 2 мы исследовали влияние длительности стимуляции, ожидая более сильных эффектов при более длительной стимуляции, наблюдаемых с помощью традиционных мер и параметров DDM.
Методы
Участники
Все методы в данном исследовании проводились в соответствии с Хельсинкской декларацией и были одобрены Комитетом по этике Исследовательского центра визуализации мозга Пекинского нормального университета. Информированное согласие было получено от всех участников до начала исследования. Каждый эксперимент включал отдельную группу участников. Все участники были правшами, имели нормальное или скорректированное до нормального зрение и не сообщали об истории неврологических или психиатрических расстройств. Априорный анализ мощности с использованием G*Power версии 3.1.9.7 [47] был проведен для определения соответствующих размеров выборки. Для Эксперимента 1 размер выборки N = 40 был рассчитан для обнаружения среднего размера эффекта (0,3) при уровне значимости α = 0,05 и мощности = 0,80 [48]. Для обеспечения достаточной мощности и учета потенциальных исключений мы набрали 48 участников для каждого из Экспериментов 1a и 1b (т.е. 24 на группу интенсивности в каждом эксперименте). Для Эксперимента 2 аналогичный анализ указал на минимальную выборку N = 17, достаточную для среднего размера эффекта при тех же статистических критериях. Для каждого из Экспериментов 2a и 2b было набрано 24 участника. Следует отметить, что участники Эксперимента 2 принадлежали к той же когорте, что и группа с высокой интенсивностью в Эксперименте 1. В частности, после завершения первого блока задачи Фланкера (при стимуляции, длившейся 10 минут), участники продолжили второй блок после продленной стимуляции (всего 20 минут). Таким образом, Эксперимент 2 использовал внутрисубъектный дизайн длительности стимуляции с использованием той же выборки. По окончании участия участники получили скромное денежное вознаграждение. Описательная статистика, включая средний возраст и стандартное отклонение для каждой группы, представлена в Таблице 1.
Материалы
Стимулы состояли из пяти горизонтально расположенных стрелок или прямых линий, с центральной (целевой) стрелкой в центре экрана. В конгруэнтных условиях все стрелки были направлены в одном направлении (например, >>>>>); в неконгруэнтных условиях стрелки-фланкеры были направлены в противоположную сторону от целевой (например, >>>>>). Нейтральные пробы имели не направленные линейные фланкеры (например, >>>>>). Задача Фланкера включала один блок из 90 проб (30 на каждое условие) в псевдослучайном порядке, с черными стимулами (RGB: 0; 0; 0) на белом фоне (RGB: 255; 255; 255).
Процедура
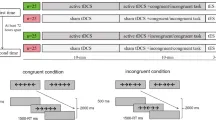
Все участники предоставили письменное информированное согласие и прошли стандартный скрининг безопасности tDCS перед экспериментом. Затем они получали 20 минут транскраниальной стимуляции постоянным током (tDCS) во время выполнения задачи Фланкера, которая использовалась для оценки когнитивного контроля. Задача проводилась в два блока, каждый из которых состоял из 90 проб и длился примерно 4-6 минут. Задача проводилась в две половины с перерывом примерно в 3 минуты между блоками. Только участники в группе с высокой интенсивностью выполняли два блока задачи Фланкера, причем первый блок начинался через 3 минуты после начала tDCS, а второй блок — через 10 минут tDCS. Никакие другие когнитивные задачи во время этого перерыва не предъявлялись, и участники оставались сидеть спокойно на протяжении всего процесса. Стимуляция начиналась за 3 минуты до начала задачи и продолжалась на протяжении всего выполнения задачи, в соответствии со стандартными протоколами. Для ясности, первый блок называется первой половиной, а второй — второй половиной, которые анализировались и сравнивались в Эксперименте 2.
Хотя все участники выполнили оба блока, в Эксперименте 1 (манипуляция интенсивностью) анализировались только данные первого блока, в то время как в Эксперименте 2 (манипуляция длительностью) в анализ были включены как первая, так и вторая половина.
Каждая проба начиналась с предъявления фиксационного креста («+») в течение 300 мс, за которым следовал пустой экран в течение 200 мс. Затем отображался стимул-фланкер и оставался на экране до тех пор, пока участник не давал ответ. После 500 мс пустого экрана начиналась следующая проба. Участникам было предложено как можно быстрее и точнее указать направление центральной целевой стрелки, используя клавишу «F» для стрелок, направленных влево, и клавишу «J» для стрелок, направленных вправо, игнорируя при этом фланкирующие стрелки по обе стороны. Стимулы предъявлялись, и ответы записывались с использованием программного обеспечения EPrime 2.0 (Psychology Software Tools, Pittsburgh, PA; https://pstnet.com/products/eprime/). В конце эксперимента участники заполнили форму обратной связи относительно своего опыта. Весь сеанс длился примерно 30 минут.
Протокол tDCS
В данном исследовании прямая стимуляция током осуществлялась с помощью стимулятора высокого разрешения (Soterix Medical) с электродами AgCl диаметром 12 мм, установленными в HDCap. Настройка высокоточной tDCS (HDtDCS) включает как аноды, так и катоды для более точной доставки тока к целевой области мозга. Для анодной HDtDCS, используемой в данном исследовании, анод использовался для повышения кортикальной возбудимости в целевой точке, в то время как катоды использовались для завершения электрической цепи, а не для производства значительных ингибирующих эффектов. Распределение симулированного электрического поля и размещение монтажа показаны на Рисунке S1 в Дополнительных материалах.
Эксперимент 1: Интенсивность стимуляции
Эксперимент 1a: Односайтовая стимуляция
Стимуляция длилась 20 минут с периодами нарастания и спада по 30 секунд. Участников просили сообщать о любом дискомфорте, но никто не просил прекратить стимуляцию. В Эксперименте 1a (односайтовая) к пре-дополнительной моторной коре (preSMA) применялись интенсивности 1 мА или 1,5 мА, с анодом, расположенным на FCz, и катодами на Cz и FC2, согласно международной системе 10–10 ЭЭГ.
Эксперимент 1b: Двухсайтовая стимуляция
Для Эксперимента 1b (двухсайтовая) стимуляция была направлена как на preSMA, так и на левый мозжечок. Один анод был расположен на FCz (с катодами на FC2 и Cz), а второй анод — на PO9 (с катодами на PO7 и O9) с интенсивностью 1 мА или 1,5 мА на анод, в соответствии с той же системой электродов.
Эксперимент 2: Длительность стимуляции
Эксперимент 2a: Односайтовая стимуляция
В Эксперименте 2a схема стимуляции была идентична Эксперименту 1a (односайтовая), но интенсивность была фиксирована на уровне 1,5 мА. Анодная стимуляция применялась над FCz, с катодами, расположенными на Cz и FC2, согласно системе 10–10 ЭЭГ. В Эксперименте 2b схема соответствовала Эксперименту 1b (двухсайтовая), также с фиксированной интенсивностью 1,5 мА на каждую точку. Аноды были расположены на FCz и PO9, с соответствующими катодами на FC2/Cz и PO7/O9 соответственно. В обоих случаях (2a и 2b) стимуляция длилась 20 минут с периодами нарастания и спада по 30 секунд. Участники выполнили два блока задачи Фланкера во время стимуляции: первый блок начался через 3 минуты (первая половина), а второй блок — через 10 минут (вторая половина).
Анализ данных
Процедуры исключения и анализа данных были одинаковыми во всех экспериментах. Для анализа включались только правильные реакции времени реакции (RT). RT короче 200 мс или длиннее 1500 мс удалялись, как и RT, превышающие 2,5 стандартных отклонения от среднего значения каждого участника. Точность была одинаково высокой во всех экспериментах (более 90%), и ошибки исключались из анализа RT. В Эксперименте 1a было удалено 1,13% проб из-за некорректных ответов, 2,08% из-за RT, превышающих 2,5 стандартных отклонения, и ни одна проба не была удалена на основе порогов абсолютного RT. В Эксперименте 1b было исключено 1,93% проб как выбросы RT, без исключения проб на основе временных порогов; уровни ошибок не сообщались отдельно. Для Эксперимента 2a было исключено 1,20% проб из-за некорректных ответов, 1,99% как выбросы и 0,02% из-за слишком короткого или слишком длинного времени. В Эксперименте 2b 1,65% проб были некорректными, 2,03% — выбросами RT, и ни одна не была удалена на основе абсолютных критериев RT. Для измерения тормозного контроля анализировались только несогласованные и нейтральные пробы [49]. Результаты, включая конгруэнтные пробы для Эксперимента 1 и Эксперимента 2, представлены на Рисунках S3 и S4 в Дополнительных материалах соответственно. Для подгонки RT и точности с помощью DDM [50] использовался набор инструментов HDDM в Python 3.8, анализирующий скорости дрейфа и время не принятия решения [20]. Для расширения основных анализов мы также включили анализ Эффекта Фланкера для каждого исследования. В частности, Эффект Фланкера рассчитывался для времени реакции, скорости дрейфа и времени не принятия решения, чтобы охватить как поведенческие, так и вычислительные индексы когнитивного контроля. Полный анализ и соответствующий рисунок представлены в Дополнительных материалах (см. Рисунок S2 в Дополнительных материалах).
Результаты
Эксперимент 1: Интенсивность стимуляции
Эксперимент 1a: Односайтовая стимуляция.
Результаты
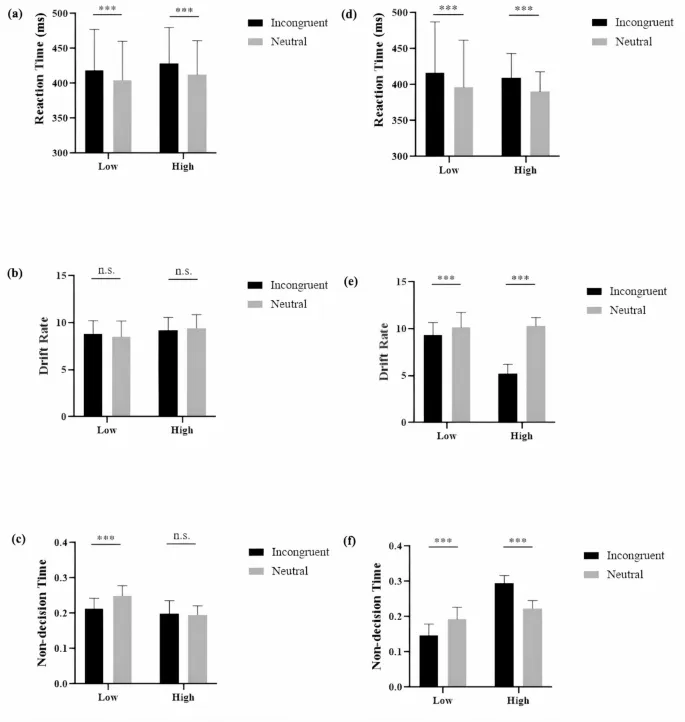
Двухфакторный дисперсионный анализ (ANOVA) для RT (Рис. 1a) показал значимый основной эффект типа пробы, F(1, 46) = 29,46, p <. 001, ηp2 = 0,39, с более медленными ответами в несогласованных (M = 423 ± 55 мс) по сравнению с нейтральными пробами (M = 408 ± 52 мс). Ни основной эффект интенсивности, ни взаимодействие не были значимыми, Fs(1, 46) < 1.
Для скорости дрейфа (Рис. 1b) не было значимых эффектов: F(1, 46) < 1 для типа пробы; F(1, 46) = 2,84, p =. 099, ηp2 = 0,06 для интенсивности, и F(1, 46) = 2,99, p =. 091, ηp2 = 0,06 для взаимодействия.
Для времени не принятия решения (Рис. 1c) результаты выявили значимый основной эффект типа пробы, F(1, 46) = 34,07, p <. 001, ηp2 = 0,43, при этом несогласованные пробы (M = 0,21 ± 0,03) давали более короткое время не принятия решения, чем нейтральные пробы (M = 0,22 ± 0,08). Результаты также показали значимый основной эффект интенсивности, F(1, 46) = 18,00, p <. 001, ηp2 = 0,28, при этом высокая интенсивность (M = 0,20 ± 0,03) давала более короткое время не принятия решения по сравнению с низкой интенсивностью (M = 0,23 ± 0,03). Взаимодействие также было значимым, F(1, 46) = 50,36, p <. 001, ηp2 = 0,52. Дальнейший анализ показал, что несогласованные пробы давали более короткое время не принятия решения, чем нейтральные пробы, только в группе с низкой интенсивностью, t(23) = 10,75, p <.001, но не в группе с высокой интенсивностью, t(23) < 1.
Двухвыборочный t-тест для Эффекта Фланкера в RT не выявил значимой разницы между группами с низкой и высокой интенсивностью.
Для скорости дрейфа двухвыборочный t-тест, сравнивающий Эффект Фланкера между группами с низкой и высокой интенсивностью, не выявил значимой разницы, t(46) = 1,73, p =.091, что указывает на то, что интенсивность стимуляции не оказала значительного влияния на разницу в накоплении информации между нейтральными и несогласованными пробами.
Для времени не принятия решения двухвыборочный t-тест показал значимую разницу между группами интенсивности, t(46) = 7,10, p <.001. В частности, при стимуляции низкой интенсивности Эффект Фланкера был отрицательным.
Эксперимент 1b: Двухсайтовая стимуляция.
Результаты
Результаты ANOVA для RT (Рис. 1d) показали значимый основной эффект типа пробы, F(1, 46) = 72,47, p <. 001, ηp2 = 0,61, с более медленными ответами в несогласованных (M = 413 ± 55 мс) по сравнению с нейтральными пробами (M = 393 ± 50 мс). Однако других значимых эффектов не было найдено, Fs(1, 46) < 1.
Для скорости дрейфа (Рис. 1e) результаты выявили значимый основной эффект типа пробы, F(1, 46) = 357,91, p <. 001, ηp2 = 0,89, при этом несогласованные пробы (M = 7,27 ± 2,42) давали более низкую скорость дрейфа, чем нейтральные пробы (M = 10,22 ± 1,30). Также был значимый основной эффект интенсивности, F(1, 46) = 96,87, p <. 001, ηp2 = 0,46, указывающий на то, что высокая интенсивность стимуляции (M = 7,74 ± 2,76) снижала скорость дрейфа по сравнению с низкой интенсивностью (M = 9,75 ± 1,51). Взаимодействие было значимым, F(1, 46) = 194,13, p <. 001, ηp2 = 0,81, и дальнейший анализ показал, что скорость дрейфа была значительно ниже для несогласованных условий по сравнению с нейтральными пробами как при низкой, t(23) = 3,99, p <.001, так и при высокой интенсивности стимуляции, t(23) = 21,03, p <.001, предполагая, что увеличение интенсивности стимуляции приводило к дальнейшему снижению скорости дрейфа для несогласованных проб по сравнению с нейтральными пробами.
Для времени не принятия решения (Рис. 1f) был найден значимый эффект типа пробы, F(1, 46) = 22,72, p <. 001, ηp2 = 0,33, с более длинным временем не принятия решения для несогласованных проб (M = 0,22 ± 0,08) по сравнению с нейтральными пробами (M = 0,21 ± 0,03). Также был значимый основной эффект интенсивности, F(1, 46) = 135,66, p <. 001, ηp2 = 0,75, при этом высокая интенсивность (M = 0,26 ± 0,04) давала более длинное время не принятия решения, чем низкая интенсивность (M = 0,17 ± 0,04). Взаимодействие было значимым, F(1, 46) = 465,74, p <. 001, ηp2 = 0,91. Дальнейший анализ показал, что несогласованные пробы имели более короткое время не принятия решения, чем нейтральные пробы, в группе с низкой интенсивностью, t(23) = 12,52, p <.001, но более длинное в группе с высокой интенсивностью, t(23) = 17,78, p <.001.
Эксперимент 2: Длительность стимуляции.
Эксперимент 2a: Односайтовая стимуляция.
Результаты
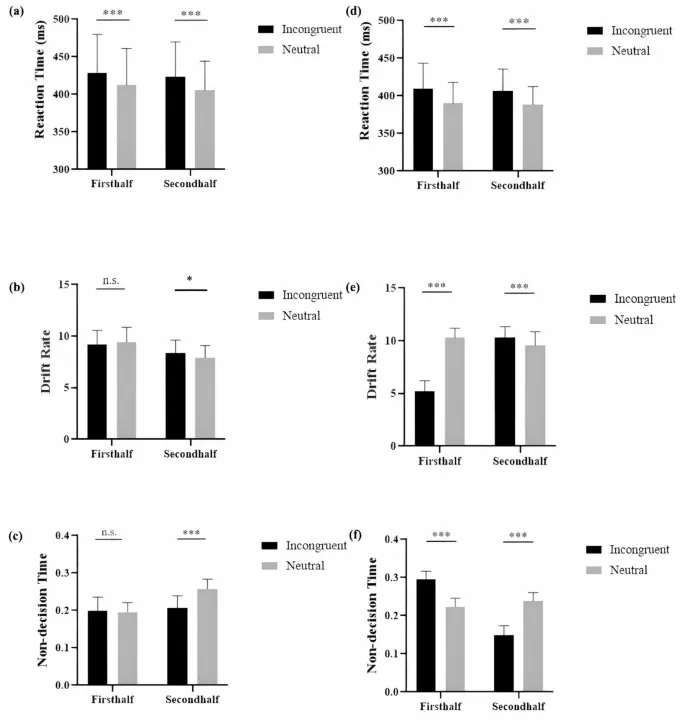
Двухфакторный ANOVA для типа пробы и длительности стимуляции по RT (Рис. 2a) показал значимый основной эффект типа пробы, F(1, 46) = 41,99, p <. 001, ηp2 = 0,48, с более медленными ответами в несогласованных пробах (M = 426 ± 48 мс) по сравнению с нейтральными пробами (M = 408 ± 44 мс). Значимых эффектов длительности или взаимодействия не было найдено, Fs(1, 46) < 1.
Для данных скорости дрейфа (Рис. 2b) наблюдался незначимый эффект типа пробы, F(1, 46) < 1, значимый основной эффект длительности, F(1, 46) = 11,71, p =. 001, ηp2 = 0,20, причем более длительная длительность (M = 8,16 ± 1,21) снижала скорость дрейфа по сравнению с первой половиной (M = 9,33 ± 1,36). Взаимодействие также было значимым, F(1, 46) = 5,81, p =. 020, ηp2 = 0,12. Дальнейший анализ показал, что разницы между несогласованными и нейтральными пробами в первой половине не было, t(23) = −1,06, p =. 302, но несогласованные пробы имели более быструю скорость дрейфа во второй половине, t(23) = 2,37, p =. 026.
Для времени не принятия решения (Рис. 2c) были найдены значимые эффекты типа пробы, F(1, 46) = 63,23, p <. 001, ηp2 = 0,58, при этом несогласованные пробы (M = 0,20 ± 0,03) давали более короткое время, чем нейтральные пробы (M = 0,23 ± 0,04). Длительность оказала значительное влияние, F(1, 46) = 18,24, p <. 001, ηp2 = 0,28, при этом вторая половина (M = 0,23 ± 0,04) имела более длительное время, чем первая половина (M = 0,20 ± 0,03). Также наблюдалось значимое взаимодействие, F(1, 46) = 82,75, p <. 001, ηp2 = 0,64, указывающее на отсутствие разницы во времени не принятия решения в первой половине, t(23) < 1, но несогласованные пробы имели более короткое время во второй половине, t(23) = 12,41, p <.001.
Эксперимент 2b: Двухсайтовая стимуляция.
Результаты
Двухфакторный ANOVA для типа пробы и длительности стимуляции по RT (Рис. 2d) показал значимый основной эффект типа пробы, F(1, 46) = 62,14, p <. 001, ηp2 = 0,58, с более медленными ответами в несогласованных пробах (M = 408 ± 31 мс) по сравнению с нейтральными пробами (M = 389 ± 25 мс). Значимого основного эффекта интенсивности стимуляции или взаимодействия не было найдено, Fs(1, 46) < 1.
Для скорости дрейфа (Рис. 2e) были найдены значимые основные эффекты для типа пробы, F(1, 46) = 188,89, p <. 001, ηp2 = 0,80, с более низкой скоростью дрейфа в несогласованных пробах (M = 7,75 ± 2,79) по сравнению с нейтральными пробами (M = 9,92 ± 1,18), и для длительности, F(1, 46) = 115,68, p <. 001, ηp2 = 0,59, причем более длительная tDCS увеличивала скорость дрейфа (M = 9,92 ± 1,18) по сравнению с короткой длительностью (M = 7,74 ± 2,76). Было найдено значимое взаимодействие, F(1, 46) = 346,44, p <. 001, ηp2 = 0,88. Скорость дрейфа была ниже для несогласованных условий в первой половине, t(23) = 21,03, p <.001, но выше во второй половине, t(23) = 3,81, p <.001.
Для времени не принятия решения (Рис. 2f) были найдены значимые основные эффекты для типа пробы, F(1, 46) = 9,25, p <. 001, ηp2 = 0,17, с более длинным временем не принятия решения в несогласованных пробах (M = 0,22 ± 0,08) по сравнению с нейтральными пробами (M = 0,21 ± 0,03), и для длительности, F(1, 46) = 110,50, p <. 001, ηp2 = 0,71, причем более длительная длительность давала более короткое время не принятия решения (M = 0,20 ± 0,05) по сравнению с короткой длительностью (M = 0,26 ± 0,04). Было найдено значимое взаимодействие, F(1, 46) = 854,39, p <. 001, ηp2 = 0,95. Время не принятия решения было более длинным для несогласованных проб в первой половине, t(23) = 17,78, p <.001, но более коротким во второй половине, t(23) = 23,86, p <.001.
Обсуждение
В Эксперименте 1a анализ RT односайтовой стимуляции выявил значимый основной эффект типа пробы, но не значимый основной эффект интенсивности стимуляции или взаимодействия. Однако последующее разложение RT в рамках DDM выявило интересные результаты. Результаты данных о скорости дрейфа показали, что увеличение интенсивности стимуляции не приводило к значительному эффекту взаимодействия, предполагая, что увеличение интенсивности не влияет на накопление информации, индексируемое скоростью дрейфа. Анализ времени не принятия решения выявил значимое взаимодействие. В частности, при стимуляции низкой интенсивности несогласованные пробы демонстрировали более короткое время не принятия решения по сравнению с нейтральными пробами, в то время как эта разница не была значимой при стимуляции высокой интенсивности. Это предполагает, что tDCS низкой интенсивности снижает перцептивные требования несогласованных условий [43,51]. В совокупности, данное исследование показало, что для односайтовой tDCS 1 мА достаточно для улучшения когнитивного контроля, и увеличение интенсивности не дает дополнительных преимуществ и может даже быть вредным.
Аналогично, при двухсайтовой стимуляции анализ RT показал значимый основной эффект типа пробы, но не значимый основной эффект или взаимодействие. Анализ DDM выявил значимое взаимодействие для скорости дрейфа: tDCS низкой интенсивности улучшала когнитивный контроль, снижая разницу в скорости дрейфа, в то время как tDCS высокой интенсивности дополнительно снижала скорость дрейфа для несогласованных проб. Анализ времени не принятия решения показал, что tDCS низкой интенсивности сокращала время для несогласованных проб, но высокая интенсивность увеличивала его, указывая на повышенный аттенционный спрос. Это предполагает, что двухсайтовая tDCS низкой интенсивности улучшает когнитивный контроль, в то время как увеличение интенсивности ухудшает его.
Было исследовано, варьируются ли эти вредные эффекты при изменении длительности стимуляции в Эксперименте 2.
Обсуждение результатов Эксперимента 2
Эксперимент 2a: Односайтовая стимуляция.
Результаты
Двухфакторный ANOVA для типа пробы и длительности стимуляции по RT (Рис. 2a) показал значимый основной эффект типа пробы, F(1, 46) = 41,99, p <. 001, ηp2 = 0,48, с более медленными ответами в несогласованных пробах (M = 426 ± 48 мс) по сравнению с нейтральными пробами (M = 408 ± 44 мс). Значимых эффектов длительности или взаимодействия не было найдено, Fs(1, 46) < 1.
Для данных скорости дрейфа (Рис. 2b) наблюдался незначимый эффект типа пробы, F(1, 46) < 1, значимый основной эффект длительности, F(1, 46) = 11,71, p =. 001, ηp2 = 0,20, причем более длительная длительность (M = 8,16 ± 1,21) снижала скорость дрейфа по сравнению с первой половиной (M = 9,33 ± 1,36). Взаимодействие также было значимым, F(1, 46) = 5,81, p =. 020, ηp2 = 0,12. Дальнейший анализ показал, что разницы между несогласованными и нейтральными пробами в первой половине не было, t(23) = −1,06, p =. 302, но несогласованные пробы имели более быструю скорость дрейфа во второй половине, t(23) = 2,37, p =. 026.
Для времени не принятия решения (Рис. 2c) были найдены значимые эффекты типа пробы, F(1, 46) = 63,23, p <. 001, ηp2 = 0,58, при этом несогласованные пробы (M = 0,20 ± 0,03) давали более короткое время, чем нейтральные пробы (M = 0,23 ± 0,04). Длительность оказала значительное влияние, F(1, 46) = 18,24, p <. 001, ηp2 = 0,28, при этом вторая половина (M = 0,23 ± 0,04) имела более длительное время, чем первая половина (M = 0,20 ± 0,03). Также наблюдалось значимое взаимодействие, F(1, 46) = 82,75, p <. 001, ηp2 = 0,64, указывающее на отсутствие разницы во времени не принятия решения в первой половине, t(23) < 1, но несогласованные пробы имели более короткое время во второй половине, t(23) = 12,41, p <.001.
Эксперимент 2b: Двухсайтовая стимуляция.
Результаты
Двухфакторный ANOVA для типа пробы и длительности стимуляции по RT (Рис. 2d) показал значимый основной эффект типа пробы, F(1, 46) = 62,14, p <. 001, ηp2 = 0,58, с более медленными ответами в несогласованных пробах (M = 408 ± 31 мс) по сравнению с нейтральными пробами (M = 389 ± 25 мс). Значимого основного эффекта интенсивности стимуляции или взаимодействия не было найдено, Fs(1, 46) < 1.
Для скорости дрейфа (Рис. 2e) были найдены значимые основные эффекты для типа пробы, F(1, 46) = 188,89, p <. 001, ηp2 = 0,80, с более низкой скоростью дрейфа в несогласованных пробах (M = 7,75 ± 2,79) по сравнению с нейтральными пробами (M = 9,92 ± 1,18), и для длительности, F(1, 46) = 115,68, p <. 001, ηp2 = 0,59, причем более длительная tDCS увеличивала скорость дрейфа (M = 9,92 ± 1,18) по сравнению с короткой длительностью (M = 7,74 ± 2,76). Было найдено значимое взаимодействие, F(1, 46) = 346,44, p <. 001, ηp2 = 0,88. Скорость дрейфа была ниже для несогласованных условий в первой половине, t(23) = 21,03, p <.001, но выше во второй половине, t(23) = 3,81, p <.001.
Для времени не принятия решения (Рис. 2f) были найдены значимые основные эффекты для типа пробы, F(1, 46) = 9,25, p <. 001, ηp2 = 0,17, с более длинным временем не принятия решения в несогласованных пробах (M = 0,22 ± 0,08) по сравнению с нейтральными пробами (M = 0,21 ± 0,03), и для длительности, F(1, 46) = 110,50, p <. 001, ηp2 = 0,71, причем более длительная длительность давала более короткое время не принятия решения (M = 0,20 ± 0,05) по сравнению с короткой длительностью (M = 0,26 ± 0,04). Было найдено значимое взаимодействие, F(1, 46) = 854,39, p <. 001, ηp2 = 0,95. Время не принятия решения было более длинным для несогласованных проб в первой половине, t(23) = 17,78, p <.001, но более коротким во второй половине, t(23) = 23,86, p <.001.
Общее обсуждение
В данном исследовании мы изучили влияние интенсивности и длительности стимуляции на поведенческую производительность как при односайтовой, так и при двухсайтовой стимуляции. Варьируя эти параметры, наши результаты выявили важное взаимодействие между параметрами tDCS, особенно с увеличением длительности. Использование как традиционного анализа RT, так и анализа DDM позволило нам количественно оценить различные когнитивные процессы в когнитивном контроле, выявив различные закономерности для односайтовой и двухсайтовой tDCS с различной интенсивностью и длительностью.
Одним из наиболее примечательных результатов этого исследования является то, что вредные эффекты, вызванные увеличением интенсивности tDCS, исчезали с увеличением длительности. В частности, это уменьшение вредных эффектов наблюдалось в группе с высокой интенсивностью (1,5 мА). Увеличение длительности tDCS улучшало скорость дрейфа и снижало время не принятия решения для несогласованных проб, которые требуют более высоких когнитивных усилий. Этот результат может отражать адаптивные изменения в функциях когнитивного контроля мозга. В частности, в Эксперименте 1 мы обнаружили, что tDCS высокой интенсивности приводила к снижению поведенческой производительности, что может указывать на чрезмерную стимуляцию определенных нейронных цепей, нарушающую нормальную когнитивную обработку и приводящую к снижению поведенческой производительности, что согласуется с существующей литературой, показывающей ограниченные дополнительные преимущества при увеличении интенсивности стимуляции [52]. Эксперимент 2 данного исследования расширил это исследование, изучив временную динамику этого вредного эффекта. Наша первоначальная гипотеза заключалась в том, что более длительная длительность приведет к более значительным эффектам. Однако результаты показали, что по мере продолжения tDCS эти вредные эффекты исчезали, что указывает на то, что мозг может задействовать компенсаторные механизмы, такие как синаптическая пластичность, в конечном итоге позволяя ему перестроиться для снижения нарушений. Насколько нам известно, данное исследование является первым, которое изучает изменения, вызванные увеличением интенсивности tDCS, с временной точки зрения. Хотя существует несоответствие между нашими первоначальными гипотезами и наблюдаемыми результатами, это, вероятно, отражает новизну применения DDM в данном исследовании. Поскольку ни одно предыдущее исследование не изучало длительность tDCS с использованием модели диффузии дрейфа (DDM), наши предсказания основывались в первую очередь на результатах исследований времени реакции, которые не отражают скрытые когнитивные компоненты, которые DDM мог бы выявить. Из-за этого мы признаем, что некоторые аспекты наших выводов являются исследовательскими и могут открыть новые направления для будущих исследований, чтобы далее изучить, как мозг реагирует на tDCS с течением времени.
В то время как предыдущие исследования (например, Jamil et al., 2017) документировали отсутствие значимых изменений в кортикальной возбудимости независимо от интенсивности tDCS (т.е. 0–2 мА) при применении tDCS к моторной коре, настоящее исследование сосредоточилось на областях мозга, участвующих в когнитивном контроле, с использованием задачи когнитивного контроля, что может способствовать этому расхождению. Возможно, что зависящие от интенсивности эффекты, наблюдаемые в когнитивных доменах, отличаются от тех, которые задокументированы в моторных областях, что может объяснить, почему мы обнаружили заметные различия между 1,0 мА и 1,5 мА в параметрах DDM. Кроме того, помимо традиционного анализа времени реакции, мы применили DDM для разложения данных времени реакции и точности, что позволило нам выявить тонкие изменения в лежащих в основе когнитивных процессах, таких как процесс накопления информации (т.е. скорость дрейфа) и процесс подготовки к движению (т.е. время не принятия решения), которые анализ RT может не выявить.
Кроме того, хотя предыдущие исследования сравнивали односайтовую и двухсайтовую tDCS, эти исследования в основном использовали кортикальную возбудимость в качестве переменной исхода [31,51,53,54,55], без дальнейшего изучения влияния этих двух парадигм стимуляции на когнитивные функции. Результаты данного исследования показали, что односайтовая tDCS в основном влияла на перцептивные процессы (время не принятия решения), в то время как двухсайтовая tDCS влияла как на перцептивные (время не принятия решения), так и на процессы накопления информации (скорость дрейфа). Это может указывать на то, что двухсайтовая tDCS может влиять на более широкий спектр нейронных цепей, потенциально усиливая связь между областями мозга. Эти результаты, в сочетании с предыдущими исследованиями [30], предполагают, что сложные когнитивные функции могут быть результатом взаимодействия между областями мозга, а не изолированной областью мозга.
Наконец, мы обнаружили, что по сравнению с традиционным анализом времени реакции и точности, модель диффузии дрейфа могла предоставить новые идеи для выявления эффекта анодной tDCS. Переводя поведенческую точность, время реакции и распределение времени реакции в скрытые когнитивные компоненты [20], DDM предлагает более полный учет процесса принятия решений. В частности, в анализе RT односайтовая и двухсайтовая tDCS демонстрировали схожие закономерности эффектов; однако благодаря разложению с помощью DDM были выявлены различные закономерности для односайтовой и двухсайтовой tDCS. Расхождение между результатами RT и результатами DDM, вероятно, связано с тем, что RT отражает сумму множественных когнитивных подпроцессов, включая накопление информации, перцептивное кодирование и моторное исполнение. Если tDCS влияет только на один из этих компонентов или влияет на несколько подпроцессов в разных направлениях, общий эффект на RT может не стать наблюдаемым, в то время как DDM способен разложить эти процессы, делая его более чувствительным к тонким изменениям [21]. Данное исследование является первым, которое использовало DDM для изучения влияния увеличения дозировки tDCS на когнитивный контроль и сложных различий между односайтовой и двухсайтовой стимуляцией, подчеркивая важность использования DDM для выявления тонких различий в эффектах tDCS на когнитивные процессы, упускаемых анализом RT.
Однако важно отметить, что наше исследование не включало плацебо-группу из-за методологических и практических ограничений. Учитывая основную цель данного исследования — одновременное сравнение нескольких параметров tDCS как в односайтовой, так и в двухсайтовой стимуляции, добавление идеально соответствующих плацебо-групп значительно увеличило бы сложность сравнений и потенциально снизило бы статистическую мощность. Таким образом, хотя отсутствие плацебо-группы ограничивает нашу способность делать окончательные выводы относительно абсолютной эффективности tDCS по сравнению с базовым уровнем, основная цель была сосредоточена на сравнении различий в эффектах стимуляции между интенсивностями, длительностью и размещением монтажа, а не на установлении причинных эффектов стимуляции по сравнению с отсутствием стимуляции. Будущие исследования, направленные на дальнейшее расширение наших выводов путем изучения абсолютных эффектов tDCS наряду с относительными эффектами варьирования параметров tDCS, выиграли бы от включения плацебо-групп в их экспериментальный дизайн.
Выводы и следствия
В заключение, впервые мы всесторонне изучили эффекты различной интенсивности и длительности tDCS как для односайтовой, так и для двухсайтовой стимуляции в рамках одного исследования, объединив как традиционные меры RT, так и фреймворк DDM. Наши выводы предполагают, что длительность стимуляции может модулировать вредные эффекты, вызванные более высокой интенсивностью tDCS, указывая на потенциальное взаимодействие между параметрами tDCS. Однако, учитывая исследовательский характер данного исследования, это взаимодействие следует интерпретировать с осторожностью и требует дальнейшего изучения. Различные результаты в отношении интенсивности и длительности tDCS подчеркивают необходимость тонкого понимания лежащих в основе механизмов tDCS для модуляции когнитивных процессов, предполагая различные закономерности для односайтовой и двухсайтовой tDCS. Согласно этим выводам, будущие исследования могут далее изучить индивидуальные различия и специфичные для задач факторы для оптимизации применения tDCS для когнитивного улучшения и терапевтических вмешательств.





