Интересное сегодня
Экспериментальные доказательства социального обучения у диких крыс Rattus norvegicus в полуприродных условиях
Животные способны приобретать информацию об окружающей среде посредством социального обучения. Социальная передача информации может влиять на скорость освоения признаков и эффективность выполнения задач. Однако часто остается неясным, как именно происходит приобретение поведенческих навыков при наличии социальной информации. В частности, роль социального обучения в освоении неинтуитивных задач остается малоизученной. Мы задались вопросом, получают ли дикие крысы Rattus norvegicus из шести полуприродных колоний пользу от взаимного обучения при освоении и выполнении неинтуитивной задачи, посредством социального обучения. Задача заключалась в открытии механизма «качели» для получения пищевого вознаграждения. Мы инициировали нововведения в четырех из шести колоний и контролировали количество обученных особей и состав по родству. Время до первого успешного манипулирования «качелями» было короче у наивных крыс, живущих с четырьмя опытными особями, по сравнению с теми, кто жил без опытных особей или с двумя опытными особями. Интервалы между успешными манипуляциями «качелями» не зависели от количества опытных особей и состава колоний по родству. Крысы в четырех колониях освоили манипулирование «качелями» путем обучения методом проб и ошибок. Наши данные показывают, что информация о решении неинтуитивной задачи по добыче пищи может передаваться социально среди диких крыс Rattus norvegicus, независимо от состава колоний по родству.
Введение
Социальное познание, включающее социальное обучение, позволяет животным приобретать и использовать информацию об окружающей среде посредством взаимодействия с социальными партнерами¹. Социальное обучение — это процесс, при котором приобретение нового поведенческого признака облегчается наблюдением за особями или взаимодействием с ними, или с их продуктами². Социальное обучение широко распространено среди различных таксонов животных¹, что способствует адаптивному принятию решений для удовлетворения экологических потребностей⁴,⁵. Социальная передача является подмножеством социального обучения, влияющим на скорость, с которой индивид (i) приобретает новый признак и (ii) демонстрирует его после приобретения². Социальная передача определяется как происходящая, когда предварительное приобретение поведенческого признака T одной особью A, выраженное либо непосредственно в выполнении T, либо в каком-либо другом поведении, связанном с T, оказывает продолжительное положительное причинно-следственное влияние на скорость, с которой другая особь B приобретает и/или выполняет T². Традиционный экспериментальный дизайн для изучения социального обучения включает пару «демонстратор-наблюдатель» с контрольными группами (например, без демонстратора)² ³. Обычно задержка приобретения способности уменьшается с увеличением числа опытных особей⁶,⁷,⁸. Однако эффект проживания с разным количеством опытных особей в естественной среде на скорость приобретения и выполнения навыков еще не оценен.
Вероятность социального обучения может быть пропорциональна коэффициенту родства между особями² ,⁹. Вертикальная передача социально усвоенных признаков от матери к потомству была продемонстрирована у приматов⁹,¹⁰,¹¹ и у американских черных медведей (Ursus americanus)¹². Однако это не наблюдалось у диких белоголовых капуцинов (Cebus capucinus) и диких мартышек верветок (Chlorocebus pygerythrus)¹³,¹⁴. Социальное обучение пищевых предпочтений у крыс Rattus norvegicus в лабораторных условиях было независимым от родства и знакомства¹⁵,¹⁶. Таким образом, существует потребность в оценке влияния родства на социальное обучение внутри групп связанных или не связанных социальных партнеров в естественных условиях.
Новое или измененное поведение, ранее не встречавшееся в популяции, является инновацией² ,¹⁷. Инновация может распространяться в популяции посредством социального и асоциального обучения¹⁷,¹⁸,¹⁹,²⁰. Социальное обучение часто инициируется инновацией одной особи путем обучения методом проб и ошибок при взаимодействии с физической средой, основываясь на личной информации²¹. Наивные особи, особенно те, кто живет без опытных, должны быть более склонны к инновациям, чем опытные особи, поскольку опытные, которые ранее приобрели признак, уже могут успешно достичь цели, в то время как наивные особи, живущие без опытных, должны изобретать новое поведение для достижения успеха.
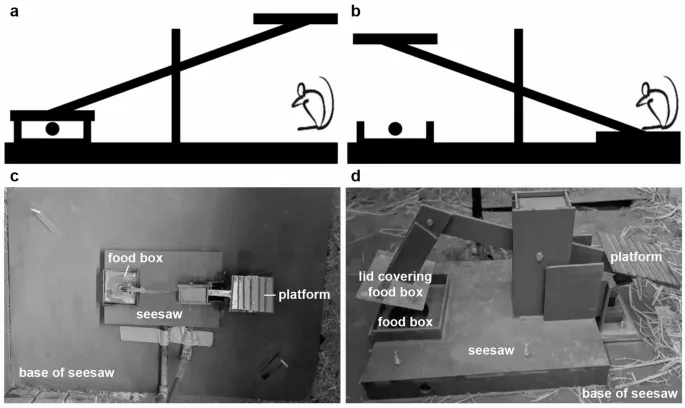
Целью данного исследования было изучить социальную передачу информации о сложной задаче посредством социального обучения у высокосоциального животного в полуприродной среде. Отдельные дикие крысы Rattus norvegicus асоциально учились открывать крышку пищевого контейнера, садясь на платформу «качелей» (Рис. 1), таким образом становясь «опытными крысами». Мы внедрили таких опытных крыс в четыре из шести колоний. Мы контролировали (i) количество особей, обученных манипулировать механизмом подачи пищи — тестом «качели»²², и (ii) состав колоний по родству. Чтобы проверить гипотезу о том, что дикие крысы Rattus norvegicus, содержащиеся в полуприродных колониях, приобретают и демонстрируют неинтуитивную задачу по добыче пищи посредством социального обучения, мы предсказали, что (1) промежуточные шаги до первого успеха (т.е. приближение к платформе или поедание пищевых вознаграждений, когда другая крыса опускает платформу, наблюдая таким образом, как конспецифические особи успешно манипулируют «качелями») будут происходить чаще у наивных крыс, живущих с большим количеством опытных особей; (2) задержка первого успешного манипулирования «качелями» будет короче у наивных крыс, живущих с большим количеством опытных особей; и (3) интервалы между успешными манипуляциями, влияющие на скорость выполнения признака после приобретения, будут короче у наивных крыс по мере увеличения числа опытных особей. Чтобы проверить гипотезу о том, что родство между особями улучшает приобретение и выполнение сложной задачи по добыче пищи в процессе социального обучения, мы предсказали, что: (4) промежуточные шаги до первого успеха будут происходить чаще, задержка первого успешного манипулирования «качелями» будет короче, и интервалы между успешными манипуляциями будут короче у наивных крыс, живущих в колониях с более высоким составом по родству; (5) мы предсказали, что вероятность успешных инновационных манипуляций будет выше у наивных крыс, чем у опытных. Крыса считалась инноватором, если она проявляла новое или измененное поведение для успешного манипулирования «качелями» (например, прямое поднятие крышки «качелей», а не сидение на платформе, которая находится на противоположном конце крышки, или сидение на платформе в колониях без опытных крыс, которые могли бы продемонстрировать это поведение), или измененное поведение (т.е. наивная или опытная крыса напрямую поднимает крышку «качелей» вместо того, чтобы сидеть на платформе в колониях с опытными крысами).
Результаты
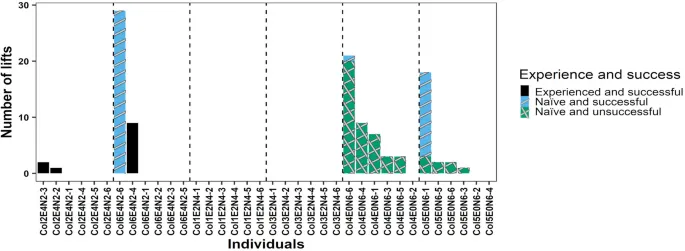
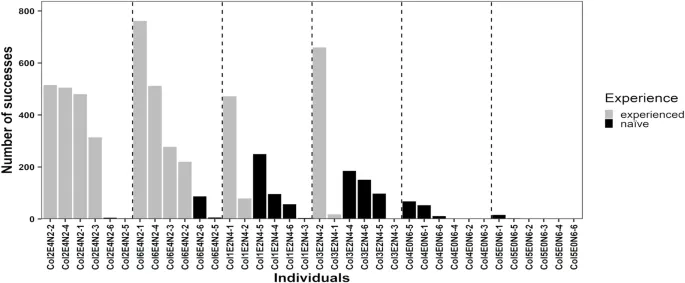
Опытные крысы были обучены выполнять неинтуитивную задачу по добыче пищи до начала основного проекта, и они освоили этот навык после 19–26 тренировочных сессий. Семнадцать из 24 наивных крыс (70,8%) успешно манипулировали «качелями» для получения пищевого вознаграждения. В эксперименте шесть из неуспешных наивных крыс жили в колониях без опытных особей, а седьмая неуспешная наивная крыса жила в колонии с 2 опытными особями (Рис. 2). Двадцать три из 24 (95,8%) наивных крыс садились на платформу, а единственная наивная крыса, которая не садилась на платформу, жила в колонии без опытных особей. Таким образом, шесть из семи неуспешных крыс манипулировали «качелями», садясь на платформу, но без доступа к пище, т.е. выполняли неэффективное манипулирование «качелями». Показатели согласованности для (i) идентификации крыс, манипулирующих «качелями», и (ii) успеха манипуляции были надежно измерены со значениями 0,999 (1605/1606 наблюдений) и 0,999 (1605/1606 наблюдений) соответственно.
Задержка первого успешного манипулирования «качелями»
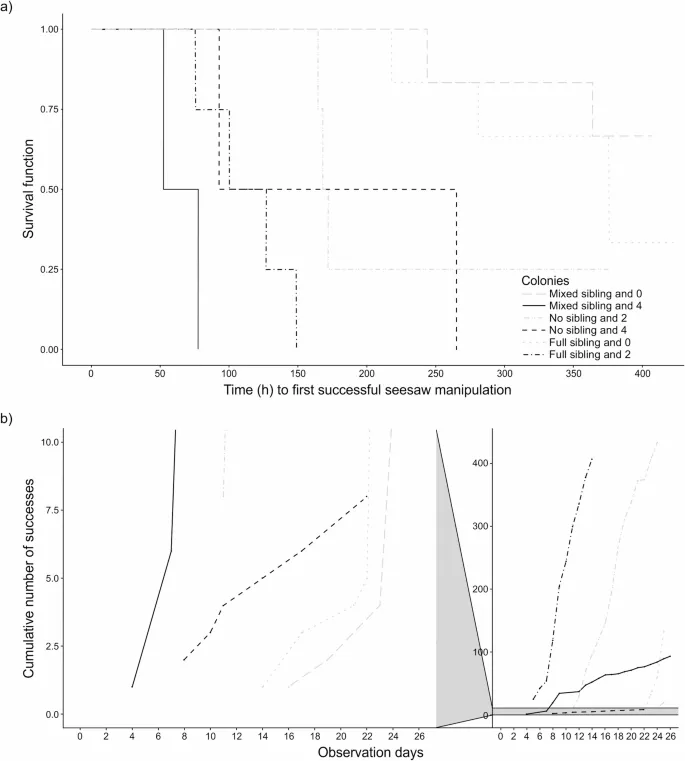
Мы провели полупараметрическую смешанную модель пропорциональных рисков Кокса для оценки влияния (i) количества опытных крыс в колонии, (ii) ее состава по родству и (iii) расположения «качелей» в вольере в качестве фиксированных эффектов на задержку наивных крыс до первого успешного манипулирования «качелями». Мы также представили полные и нулевые сравнения. Полная модель лучше подходила для данных, чем нулевая (только интерцепт) модель (Χ² = 14,4, p = 0,006). Задержка первого успешного манипулирования «качелями» была короче у наивных крыс, живущих с четырьмя опытными особями, по сравнению с наивными крысами, живущими без опытных особей (4 против 0 опытных особей: p = 0,025, Таблица 1; Рис. 3a) и у наивных крыс, живущих с четырьмя опытными особями, по сравнению с наивными крысами, живущими с двумя опытными особями (4 против 2 опытных особей, p = 0,006, Таблица S1, Рис. 3a). Однако значительной разницы в задержке первого успешного манипулирования «качелями» у наивных крыс, живущих с двумя опытными особями, по сравнению с наивными крысами, живущими без опытных особей, не было (2 против 0 опытных особей: p = 0,24, Таблица 1; Рис. 3a). Эти результаты частично подтвердили наше направленное предсказание относительно приобретения успешного манипулирования «качелями» по мере увеличения числа опытных особей.
Задержка первого успешного манипулирования «качелями» наивными крысами была короче в условиях отсутствия родственных связей между сиблингами, чем в условиях смешанных родственных связей сиблингов (без сиблингов против смешанных сиблингов: p = 0,024, Таблица 1; Рис. 3a). Задержка первого успешного манипулирования «качелями» наивными крысами не отличалась между полными родственными связями сиблингов и смешанными родственными связями сиблингов (полные сиблинги против смешанных сиблингов: p = 0,30, Таблица 1; Рис. 3a), а также между полным родством сиблингов и отсутствием родства сиблингов (полные сиблинги против отсутствия сиблингов: p = 0,12, Таблица S1, Рис. 3a). Эти результаты не соответствовали нашим направленным предсказаниям относительно влияния родства на задержку освоения задачи. Задержка первого успешного манипулирования «качелями» наивными крысами была короче после изменения расположения «качелей», чем в их первоначальном расположении (измененное положение против первоначального положения: p = 0,02, Таблица 1).
Доверительные интервалы некоторых оцененных параметров были довольно широкими, что указывает на неточность в оценках. Тем не менее, мы представили полные и нулевые сравнения моделей, чтобы избежать завышенных ошибок первого рода, вероятных ошибок первого рода и значений стабильности модели.
Промежуточные шаги к освоению успешного манипулирования «качелями»
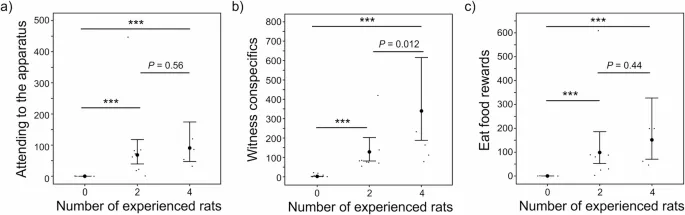
Полные модели лучше подходили, чем нулевые (только интерцепт) модели (т.е. приближение к платформе: Χ² = 26,51, p < 0,001; наблюдение за конспецифическими особями: Χ² = 27,59, p < 0,001; поедание пищевых вознаграждений: Χ² = 25,89, p < 0,001). До первого успешного манипулирования «качелями» наивные крысы в колониях с 2 или 4 опытными особями (i) чаще приближались к аппарату, когда другая крыса опускала платформу (4 против 0: p < 0,001; 2 против 0: p < 0,001, Таблица 1; Рис. 4a), (ii) чаще наблюдали, как конспецифические особи успешно манипулируют «качелями» (4 против 0: p < 0,001; 2 против 0: p < 0,001, Таблица 1; Рис. 4b), и (iii) чаще ели, когда другая крыса опускала платформу (4 против 0: p < 0,001; 2 против 0: p < 0,001, Таблица 1; Рис. 4c), по сравнению с наивными крысами в колониях без опытных особей. Наивные крысы в колониях с 4 опытными особями чаще наблюдали, как конспецифические особи успешно манипулируют «качелями», чем наивные крысы в колониях с 2 опытными особями (4 против 2: Оценка ± SE = 0,97 ± 0,39, p = 0,012; Рис. 4b, Таблица S1). Не было различий между наивными крысами в колониях с 4 или 2 опытными особями по частоте, с которой они (i) приближались к аппарату до первого успешного манипулирования, когда другая крыса опускала платформу (4 против 2: Оценка ± SE = 0,28 ± 0,48, p = 0,56; Рис. 4a, Таблица S1) и (ii) ели до первого успешного манипулирования, когда другая крыса опускала платформу (4 против 2: Оценка ± SE = 0,43 ± 0,56, p = 0,44; Рис. 4c, Таблица S1).
Наивные крысы в колониях с полным родством сиблингов реже приближались к аппарату, чем наивные крысы в колониях без сиблингов (полные сиблинги против отсутствия сиблингов: Оценка ± SE = -1,12 ± 0,40, p = 0,005, Таблица S1). В остальном состав колоний по родству не влиял на частоту, с которой наивные крысы (i) приближались к аппарату до первого успешного манипулирования, когда другая крыса опускала платформу (полные сиблинги против смешанных сиблингов: p = 0,17; отсутствие сиблингов против смешанных сиблингов: p = 0,69, Таблица 1), (ii) наблюдали, как конспецифические особи успешно манипулируют «качелями» (полные сиблинги против смешанных сиблингов: p = 0,17; отсутствие сиблингов против смешанных сиблингов: p = 0,61, Таблица 1; полные сиблинги против отсутствия сиблингов: Оценка ± SE = 0,30 ± 0,34, p = 0,38, Таблица S1) и (iii) ели до первого успешного манипулирования, когда другая крыса опускала платформу (полные сиблинги против смешанных сиблингов: p = 0,21; отсутствие сиблингов против смешанных сиблингов: p = 0,85, Таблица 1; полные сиблинги против отсутствия сиблингов: Оценка ± SE = -0,86 ± 0,46, p = 0,064, Таблицы S1). Эти результаты не соответствовали нашему направленному предсказанию о влиянии состава по родству на промежуточные шаги к освоению успешного манипулирования «качелями».
Скорость выполнения манипуляций с «качелями» после приобретения навыка
Сравнение полной и нулевой модели показало, что полная модель не лучше соответствовала данным, чем нулевая (только интерцепт) модель (Χ² = 8,63, p = 0,07), поэтому любые значимые эффекты, вероятно, являются ошибками первого рода²³. Интервалы между успешными манипуляциями «качелями» были короче у наивных крыс, живущих с четырьмя опытными особями, по сравнению с наивными крысами, живущими без опытных особей (4 против 0: p = 0,03, Таблица 1; Рис. 3b, однако это, вероятно, ошибка первого рода) и у наивных крыс, живущих с четырьмя опытными особями, по сравнению с наивными крысами, живущими с двумя опытными особями (4 против 2: Оценка ± SE = -2,03 ± 0,83, p = 0,01, однако это, вероятно, ошибка первого рода, Таблица S1). Интервалы между успешными манипуляциями «качелями» не отличались между наивными крысами, живущими с двумя опытными особями, по сравнению с наивными крысами, живущими без опытных особей (2 против 0: p = 0,88, Таблица 1; Рис. 3b). Другими словами, количество опытных особей не оказало явного влияния на скорость выполнения манипуляций с «качелями» после приобретения навыка. Интервалы между успешными манипуляциями «качелями» не отличались между составами по родству (полные сиблинги против смешанных сиблингов: p = 0,10; отсутствие сиблингов против смешанных сиблингов: p = 0,08, Таблица 1; полные сиблинги против отсутствия сиблингов: Оценка ± SE = -0,22 ± 0,78, p = 0,78, Таблица S1). Изменение расположения «качелей» не повлияло на интервалы между успешными манипуляциями «качелями» (измененное положение против первоначального положения: p = 0,73, Таблица 1).
Инновации
Крысы продемонстрировали новое или измененное поведение для управления «качелями» в 4 из 6 колоний (см. Дополнительную информацию, S3). Наивные крысы, живущие в колониях без опытных особей, освоили манипулирование «качелями» путем сидения на платформе (Рис. 2 и 3) или путем прямого поднятия крышки без использования механизма «качелей» путем сидения на платформе (Рис. 5). Наивные и опытные крысы в двух других колониях (колонии 2 и 6, обе с 2 опытными и 4 наивными крысами) успешно манипулировали «качелями», напрямую поднимая крышку для получения пищевого вознаграждения, чего раньше не демонстрировала ни одна опытная крыса (Рис. 5). Тринадцать из 36 крыс в общей сложности (36,1%) пытались напрямую поднять крышку, чтобы манипулировать «качелями», вместо того чтобы садиться на платформу для манипулирования «качелями». Только 6 крыс успешно манипулировали «качелями», напрямую поднимая крышку. Прямое поднятие крышки без сидения на платформе для манипулирования «качелями» составило менее 1% от всех успешных манипуляций «качелями» в нашем исследовании (57 из 5909 успешных манипуляций, т.е. 0,96%). Среднее количество попыток поднять крышку на одну крысу, исключая крыс, которые не пытались поднять, составило 5,36 ± 2,72. Сравнение полной и нулевой модели показало, что полная модель не лучше соответствовала данным, чем нулевая (только интерцепт) модель (Χ² = 4,82, p = 0,09). Вероятность успешных манипуляций «качелями» с использованием инновационных манипуляций (т.е. прямое поднятие крышки или сидение на платформе для крыс, живущих в колониях без опытных особей; прямое поднятие крышки для крыс, живущих в колониях с опытными особями) не отличалась между наивными и опытными крысами (наивные крысы против опытных крыс: p = 0,76, Таблица 1).
Обсуждение
Наши результаты предоставляют доказательства, соответствующие тому, что социальная передача информации влияет на скорость приобретения, но не на скорость выполнения, неинтуитивной задачи по добыче пищи посредством социального обучения у диких крыс Rattus norvegicus, живущих в полуприродных колониях. Результаты частично подтвердили наше направленное предсказание о том, что задержка первого успешного манипулирования «качелями» должна уменьшаться по мере увеличения числа опытных крыс в колонии. Наивные крысы, живущие без опытных особей, по-видимому, сначала нуждались в асоциальных обучающихся, чтобы освоить навык путем проб и ошибок при взаимодействии с физической средой, за чем последовала социальная передача информации внутри их колоний. Наши результаты согласуются с предыдущими лабораторными исследованиями, которые сообщали об уменьшении задержки в освоении пищевых предпочтений у крыс Rattus norvegicus⁷, пищевого поведения у голубей (Columba livia)⁶ и предпочтений в выборе мест кормления у гуппи (Poecilia reticulata)⁸ по мере увеличения числа присутствующих демонстраторов. В предыдущих исследованиях наивные особи не жили и не взаимодействовали с экспериментально манипулируемым количеством демонстраторов на протяжении всего исследования.
Особи, которые учатся социально, являются «искателями информации», тогда как особи, которые учатся асоциально, являются «производителями информации»²⁴,²⁵. Асоциальные обучающиеся могут нести большие временные и энергетические затраты при взаимодействии со средой по сравнению с социальными обучающимися, для которых приобретение информации путем наблюдения за другими кажется недорогим²⁶. Наивные крысы, живущие с 2 и 4 опытными особями, имели доступ к дополнительной социальной информации, т.е. приближались к аппарату, наблюдали за успешным манипулированием «качелями» конспецифическими особями и ели пищевые вознаграждения до первого успешного манипулирования «качелями», чаще, чем наивные крысы, живущие без опытных особей, что может объяснить более короткие задержки до первого успешного манипулирования «качелями» у наивных крыс, живущих с 2 или 4 опытными особями, по сравнению с наивными крысами, живущими без опытных особей. Наивные крысы, живущие с 4 опытными особями, чаще наблюдали за успешным манипулированием «качелями» конспецифическими особями, чем наивные крысы, живущие с 2 опытными особями. Опытные крысы, живущие в двух колониях с 4 опытными особями, успешно манипулировали «качелями» 1814 и 1772 раза соответственно, в то время как опытные крысы, живущие в двух колониях с 2 опытными особями, успешно манипулировали «качелями» 551 и 678 раз (Рис. 2). Таким образом, наивные крысы, живущие с 4 опытными особями, имели больше возможностей наблюдать за успешным манипулированием «качелями» опытными крысами, чем наивные крысы, живущие с 2 опытными особями, что, вероятно, объясняет, почему задержка первого успешного манипулирования «качелями» была короче у наивных крыс, живущих с 4, чем с 2 опытными особями. Согласуясь с этими аргументами, в предыдущих лабораторных исследованиях наблюдение за большим количеством демонстраторов, без взаимодействия с демонстраторами, сокращало задержку освоения признаков у наивных особей⁶,⁷,⁸, а скорость приобретения была ниже у наивных субъектов без демонстраторов, чем у субъектов с демонстраторами²⁷,²⁸,²⁹.
Предыдущие исследования показали, что социальная передача пищевых предпочтений у диких крыс Rattus norvegicus, содержащихся в лабораторных условиях, была независима от родства и знакомства¹⁵,¹⁶, а неродственные самцы диких крыс чаще отвечали взаимностью на полученное кормление, чем родные братья³⁰. В контролируемом лабораторном эксперименте самки диких крыс Rattus norvegicus обменивались пищей с социальными партнерами, применяя правило принятия решений прямой взаимности, которое гласит «помоги тому, кто помог тебе», а не копируя путем имитации³¹. Согласуясь с предыдущими выводами о социальном обучении у крыс Rattus norvegicus¹⁵,¹⁶, наши направленные предсказания о влиянии состава колоний по родству на склонность к социальному обучению экспериментальных субъектов не подтвердились.
Мы обнаружили свидетельства инноваций в виде нового или измененного поведения, ранее не наблюдавшегося в четырех из шести колоний. Особи могут приобретать информацию об окружающей среде, используя обучение методом проб и ошибок при взаимодействии с средой, т.е. используя личную информацию²¹. Инноваторы ввели поднятие крышки напрямую для манипулирования «качелями» без сидения на платформе, что начало распространяться внутри колоний до конца исследования. Прямое поднятие крышки могло бы распространиться на большее число членов колонии, если бы крысы изобрели эту практику раньше, чтобы их сородичи имели больше времени для получения социальной информации и изучения этого альтернативного способа получения пищи. Это предположение частично подтверждается наблюдением, что через 1 месяц во время тренировочных сессий для последующего исследования кооперации (неопубликованные данные) еще 15 крыс поднимали крышку напрямую для доступа к пище.
Ограничением данного исследования является то, что доверительные интервалы для некоторых оцененных параметров были довольно широкими, что отражает неточность в оценках. Однако мы представили полные и нулевые сравнения моделей, чтобы избежать завышенных ошибок первого рода. Размер выборки наивных крыс Rattus norvegicus в нашем исследовании составил 24 особи, распределенные по шести колониям. В будущем было бы целесообразно повторить это исследование с большим размером выборки.
Мы не ставили целью определить точные механизмы социального обучения у крыс Rattus norvegicus, которые приобрели и выполняли навык после его приобретения, поэтому мы не можем различить различные процессы социального обучения. Торп³² предположил, что простейшие процессы социального обучения, такие как локальное усиление (local enhancement), действовали в диких популяциях. В нашем исследовании задержка первого успешного манипулирования «качелями» увеличилась после изменения расположения «качелей», что может быть более вероятно объяснено тем, что больше крыс освоили навык позже в исследовании, чем до изменения расположения, чем локальным усилением. В предыдущих исследованиях экскреторные метки демонстраторов влияли на социальную передачу пищевых предпочтений у крыс Rattus norvegicus³³,³⁴. Наши опытные крысы жили с наивными крысами почти 1 месяц, и потенциальные экскреторные метки опытных крыс не удалялись после каждого манипулирования «качелями» опытной крысой. Таким образом, стимуляционное усиление (stimulus enhancement) является одним из возможных механизмов социального обучения, который мог повлиять на наши результаты. Для проверки действующих механизмов социального обучения, таких как локальное усиление и стимуляционное усиление, потребуется серия экспериментов, которые было бы легче провести с использованием экспериментальных лабораторных методов, чем изучать полуприродные колонии².
Данное исследование подтвердило гипотезу социального обучения и показало, что социальная информация для неинтуитивной задачи по добыче пищи может передаваться социально через полуприродные колонии диких крыс Rattus norvegicus. Наивные крысы успешно манипулировали «качелями» быстрее, когда жили с четырьмя опытными особями, по сравнению с жизнью (i) с двумя опытными особями или (ii) без опытных особей. Задержка успешного манипулирования «качелями» не уменьшалась с увеличением состава по родству, однако наивные крысы, живущие без сиблингов, осваивали «качели» быстрее, чем наивные крысы, живущие со смесью сиблингов и не-сиблингов. Количество раз, когда наивные крысы находились рядом с «качелями» во время манипулирования ими другой крысой, т.е. наблюдали за манипулированием «качелями» конспецифическими особями, и ели пищевые вознаграждения, когда другая крыса манипулировала «качелями», до собственного приобретения навыка наивными крысами, являлись промежуточными шагами к социальному обучению, подтверждая, что наивные крысы получали информацию от конспецифических особей до приобретения навыка. Интервалы между успешными манипуляциями не зависели от количества опытных особей и состава колоний по родству.
Методы
Модельная система
Знания о поведении диких крыс Rattus norvegicus довольно ограничены³⁵,³⁶,³⁷. Размер колонии может достигать более 150 особей³⁶. Естественные колонии организованы в переменные подгруппы, от одиночных особей обоих полов до пар, однополых групп и гаремов с потомством и без него³⁵,³⁷. Крысы Rattus norvegicus, пойманные в 9 местах, имели низкую генетическую родство между особями, пойманными в тех же местах, но демонстрировали высокие уровни генетического разнообразия и генетического структурирования на небольших географических расстояниях³⁸.
Полевые исследования крыс Rattus norvegicus, в которых неформально сообщалось о явлениях, похожих на социальную передачу пищевого поведения и предпочтений³⁵,³⁹, впоследствии были подтверждены лабораторными исследованиями, проведенными с инбредными лабораторными линиями крыс Rattus norvegicus²⁷,⁴⁰,⁴¹ и дикими крысами Rattus norvegicus⁴². Молодые крысы Rattus norvegicus учились у конспецифических особей, где, когда и что есть⁴², в то время как взрослые самцы крыс Rattus norvegicus осваивали пищевые предпочтения по экскреторным меткам и вкусовым сигналам конспецифических особей³³,³⁴. Кроме того, взрослые самцы крыс Rattus norvegicus улучшали свою эффективность поиска пищи в присутствии обученных демонстраторов с доступной пищей, и они демонстрировали более короткую задержку начала раскопок и большее количество выкопанных пищевых предметов в этих условиях²⁷.
Крысы Rattus norvegicus — высокосоциальные животные⁴³,⁴⁵, которые могут различать родственников и неродственников⁴⁴,⁴⁵, различные степени родства⁴⁶, членов колонии и нарушителей⁴⁷, а также отдельных особей (т.е. истинное индивидуальное распознавание⁴⁸). Мы использовали крыс Rattus norvegicus, известных своей способностью к сотрудничеству, используя детальную информацию от своих социальных партнеров³⁰,⁴⁹,⁵⁰,⁵¹,⁵²,⁵³,⁵⁴. Крысы Rattus norvegicus учитывают потребности своих партнеров⁵⁰,⁵¹,⁵⁵, их просьбы⁵⁶,⁵⁷,⁵⁸ и их полезность⁵²,⁵⁴,⁵⁹,⁶⁰,⁶¹ при сотрудничестве. Эти и другие исследования иллюстрируют способность крыс Rattus norvegicus адекватно реагировать на социальные сигналы⁶². Большинство исследований крыс Rattus norvegicus проводилось в лаборатории. Для изучения поведения крыс в полуприродных условиях мы создали шесть колоний диких крыс Rattus norvegicus в наружных вольерах.
Экспериментальные животные и условия содержания
Пятьдесят шесть аутбредных самок диких крыс Rattus norvegicus (источник: Behavioural Physiology Unit, Groningen Institute of Evolutionary Life Sciences, University of Groningen, The Netherlands) были доставлены на Этиологическую станцию Хасли Бернского университета, Швейцария. Крысы были индивидуально маркированы белым красителем для шерсти (крысы были привычны к запаху и применению) и проколами ушей. Узоры окрашивания шерсти позволяли нам идентифицировать каждую отдельную крысу в колонии. Если после прокола уха появлялась кровь, мы останавливали ее, мягко прижимая ухо бумажной салфеткой в течение 10 секунд. Крысы содержались в помещении группами по 4–6 сиблингов в каждой клетке для содержания (80 см x 50 см x 37,5 см). Чтобы избежать внутривидовой конкуренции за самок и возможной гибели, вызванной открытой агрессией, только самки были включены в формирование каждой колонии. Крысы были привычны к обращению (см. Дополнительную информацию, S2 для более подробной информации). Исследование проводится в соответствии с рекомендациями ARRIVE. Лицензия на проведение экспериментов с животными была выдана Швейцарским федеральным ветеринарным управлением кантона Берн (номер лицензии BE 55/18) М.Т. Разрешение на проведение необходимых исследований было выдано Бернским университетом (номер разрешения EAC201216T#212) М.Т. Эксперимент проводился в соответствии с соответствующими руководящими принципами и нормами.
После 6 недель акклиматизации, т.е. постепенного снижения температуры, крысы были перемещены во внешние вольеры в полуприродные условия. Каждый вольер (294 см x 208 см x 258 см) состоял из (i) цементного пола, покрытого 5 см грунта, и стенок из нержавеющей стали, (ii) зоны грунта (132 см x 105 см и глубиной 40 см) для рытья и строительства туннелей, (iii) 3 деревянных укрытий, (iv) 3 ламп обогрева (включались, когда температура была < 6 °C), (v) 2 ПВХ-труб, (vi) 2 кусков дерева, (vii) 4 инфракрасных лампочек (Дополнительная информация, Рис. S1) и (viii) Raspberry Pi Model 3 B+ с камерой Raspberry Pi H с объективом «рыбий глаз» и ночным видением для записи видео с частотой 30 кадров/с и разрешением 1024 пикселей по ширине и 768 пикселей по высоте. Сено и солома предоставлялись еженедельно для строительства гнезд. Зерновая смесь дополнительно предоставлялась пять раз в неделю, а свежие фрукты или овощи — два раза в неделю. Мы проводили ежедневные, еженедельные и ежемесячные медицинские осмотры.
Аппарат
Мы предоставили 1 «качели» на каждую колонию. Каждые «качели» состояли из платформы, соединенной рычагом с крышкой, которая закрывала пищевые вознаграждения в кормушке. «Качели» стояли на ПВХ-основании (102 см x 72 см x 0,5 см, Рис. 1a и c). Крысы могли опускать платформу, садясь на нее, но также могли получить доступ к пище, поднимая закрывающую крышку (Рис. 1b и d). Чтобы отговорить крыс от доступа к пищевым вознаграждениям путем поднятия крышки, она была окружена 4 кусками ПВХ, что затрудняло поднятие крышки. Похожие «качели» использовались для изучения кооперации у кеа (Nestor notabilis)²². Когда крыса опускала платформу, это соединялось с электромагнитом и двумя микропереключателями (Рис. 1d). Электромагнит соединялся с микропереключателем, источником питания 24 В и реле времени, которое удерживало лоток для пищи открытым в течение 63 секунд. Второй микропереключатель соединялся с Raspberry Pi Model 3 B+, который автоматически регистрировал даты и время открытия и закрытия «качелей». Данные передавались с Raspberry Pi на сервер по Ethernet-кабелю.
Основное исследование: обучение
В пилотном исследовании мы использовали 20 крыс (см. Дополнительную информацию, S3), а в основном исследовании тестировали 36 крыс. Для экспериментального инициирования манипуляций «качелями» 12 крыс были случайным образом отобраны из разных семей для обучения. Каждая крыса помещалась в экспериментальную клетку (80 см x 50 см x 37,5 см) с «качелями» в тренировочной комнате и возвращалась в свою клетку для содержания после каждой тренировочной сессии. Успешная манипуляция «качелями» определялась как опускание платформы путем сидения на платформе или поднятие крышки и поедание пищевого вознаграждения. Неуспешная манипуляция «качелями» определялась как: 1) неполное опускание платформы до соприкосновения с микропереключателями и электромагнитом, или ii) опускание платформы без поедания пищевого вознаграждения. Пищевыми вознаграждениями были половинки арахиса в течение первых 5 успешных сессий для повышения мотивации манипулировать «качелями», а затем они были заменены овсом. Наблюдатель записывал количество успешных и неуспешных манипуляций. Критерием для того, чтобы считать крысу обученной, далее «опытными крысами», было ≥ 4 успешных манипуляций за 15-минутную сессию в течение 2 последовательных дней (критерий был достигнут после 19–26 тренировочных сессий). Обученные крысы успешно манипулировали «качелями», садясь на платформу, а не пытаясь поднять крышку.
Основное исследование: процедура
Мы экспериментально инициировали манипуляции «качелями» в 4 из 6 колоний и манипулировали количеством опытных особей и составом колоний по родству (6 крыс/колония). У крыс был 1 неделя для акклиматизации к новой физической и социальной среде, а средняя масса и возраст крыс составляли 193 г ± 5 г и 72 дня ± 0,5 дня соответственно. В 2 колониях было 4 опытные крысы, в 2 колониях — 2 опытные крысы, и в 2 колониях — 0 опытных крыс (Таблица 2). В 2 колониях были полные сиблинги (все сестры), в 2 колониях — без сиблингов (без сестер), и в 2 колониях — смешанные сиблинги (3 пары сестер) (Таблица 2). Наивные крысы не имели предыдущего опыта работы с «качелями». Исследование длилось 26 дней наблюдений с 28 ноября 2019 года по 31 декабря 2019 года и yielded 2195,42 часа видеозаписей. Исследование проводилось при красном освещении ночью, с ежедневными температурами от 0 °C до 8 °C.
«Качели» закрывались клетчатой крышкой каждое утро, чтобы предотвратить доступ, и открывались вечером, чтобы крысы имели доступ к ним ночью во время своей активной фазы. Кормовой ящик был наполнен овсом в качестве пищевых вознаграждений. Чтобы учесть локальное усиление, положение «качелей» было изменено путем перемещения «качелей» в другое место внутри вольеров через первые 15 дней (Дополнительная информация, Рис. S1a и Рис. S1b). Исследователи просматривали видео и записывали идентификацию крыс, которые (i) опускали платформу, (ii) наблюдали за успешным манипулированием «качелями» конспецифическими особями из любого места в вольере, кроме как в кормовой клетке, укрытиях или зоне с грунтом (Рис. S1), (iii) приближались к аппарату, т.е. находились на основании аппарата, когда им манипулировал конспецифик, как прокси-индикатор наблюдения за манипулированием «качелями» конспецифическими особями, (iv) ели пищевое вознаграждение, и (v) манипулировали «качелями» новым или измененным способом (инновация), например, поднимая крышку вместо сидения на платформе. Для каждой наивной крысы задержка первого успешного манипулирования «качелями» записывалась как время от начала эксперимента в колонии до первого успешного манипулирования «качелями». Если наивная крыса вообще не смогла успешно манипулировать «качелями» в течение исследования, эта задержка определялась как время от начала эксперимента в колонии до конца исследования, и мы записывали, что событие не произошло. Для каждой наивной крысы, которая впоследствии освоила успешное манипулирование «качелями», мы записывали интервалы между успешными манипуляциями «качелями», начиная с интервала между 1-м и 2-м успехами, затем между 2-м и 3-м успехами и так далее, до конца исследования. Записывалось количество успешных и неуспешных манипуляций «качелями».
Статистический анализ
Для оценки надежности между наблюдателями мы рассчитали индекс согласованности между 2 наблюдателями для идентификации крыс, манипулирующих «качелями», и для определения того, был ли манипуляция успешной⁶³. Для исследования влияния на задержки наивных крыс до первого успешного манипулирования «качелями», т.е. приобретения нового поведенческого признака, мы провели полупараметрическую смешанную модель пропорциональных рисков Кокса для оценки влияния (i) количества опытных крыс в колонии, (ii) ее состава по родству и (iii) расположения «качелей» в вольере как зависящего от времени ковариата в качестве фиксированных эффектов на задержку наивных крыс до первого успешного манипулирования «качелями». Номер колонии был включен в качестве случайного эффекта перехвата, и не было теоретически важных случайных наклонов. Мы сравнили полную модель с нулевой моделью. Допущение пропорциональных рисков было выполнено⁶⁴.
Влияние количества опытных крыс и состава колоний по родству на задержку первого успешного манипулирования «качелями» может быть объяснено возникновением потенциальных промежуточных шагов в приобретении социальной информации до первого успешного манипулирования «качелями». Мы оценили, повлияли ли промежуточные шаги на приобретение первого успешного манипулирования «качелями», и каждая модель является прокси-показателем наблюдения наивными крысами за манипулированием «качелями» конспецифическими особями. Для оценки влияния количества опытных крыс и состава колоний по родству на количество раз, когда каждая наивная крыса приближалась к аппарату, когда другая крыса опускала платформу, до первого успеха, была проведена обобщенная линейная смешанная модель с квазипуассоновским (пуассоновским логнормальным) распределением. Для оценки влияния количества опытных крыс и состава колоний по родству на совокупное количество раз, когда каждая наивная крыса наблюдала, как другая крыса успешно манипулирует «качелями», до первого успеха, была проведена обобщенная линейная смешанная модель с квазипуассоновским (пуассоновским логнормальным) распределением. Для оценки влияния количества опытных крыс и состава колоний по родству на совокупное количество раз, когда каждая наивная крыса ела вознаграждение, когда другая крыса опускала платформу, до первого успеха, была проведена обобщенная линейная смешанная модель с квазипуассоновским (пуассоновским логнормальным) распределением. Для этих моделей колонии и значение уровня наблюдения⁶⁵ были случайными эффектами перехвата, и мы применили смещение для учета задержки каждой крысы до первого успешного манипулирования «качелями» или до конца исследования для крыс, которые не освоили задачу. Мы учли все теоретически важные случайные наклоны. Мы сравнили полные модели с нулевыми.
Для проверки влияния на интервалы между успешными манипуляциями «качелями», т.е. на скорость выполнения приобретенного навыка, мы провели параметрический анализ истории событий с распределением Вейбулла, поскольку допущение пропорциональных рисков не было выполнено. Фиксированными эффектами были (i) количество опытных крыс в колонии, (ii) ее состав по родству и (iii) расположение «качелей» в вольере как зависящий от времени ковариат, и мы включили идентификаторы крыс и колонии в качестве случайных эффектов, т.е. общий гамма-фрайлти. Мы сравнили полную модель с нулевой моделью, т.е. без количества опытных крыс и состава по родству.
Линейная смешанная модель с гауссовым распределением была проведена для оценки влияния опыта предыдущей крысы (наивная против опытной) и родства крыс (сестра против не-сестры) в качестве фиксированных эффектов на задержку наивных крыс до успешного манипулирования «качелями» после того, как ими манипулировала другая крыса. Колонии и отдельные крысы были случайными эффектами перехвата. Остатки модели были нормально распределены, когда мы логарифмически преобразовали зависимую переменную, т.е. задержку наивных крыс до успешного манипулирования «качелями» после того, как ими манипулировала другая крыса. Мы учли все теоретически важные случайные наклоны. Мы сравнили полную модель с нулевой моделью.
Была проведена обобщенная линейная смешанная модель с биномиальным распределением для оценки опыта крыс, т.е. наивных или опытных, на вероятность успешных манипуляций «качелями» с использованием инновационных манипуляций (т.е. поднятие крышки и сидение на платформе для крыс, живущих в колониях без опытных особей; поднятие крышки для крыс, живущих в колониях с опытными особями). Идентификаторы крыс и колонии были случайными эффектами перехвата. Поскольку было мало успешных открытий «качелей» с использованием инноваций, мы включили только опыт крыс, т.е. наивных или опытных, в качестве фиксированного эффекта. Мы сравнили полную модель с нулевой моделью.
Чтобы проверить значимость интересующих нас фиксированных эффектов, мы провели полные и нулевые сравнения моделей, чтобы избежать скрытого множественного тестирования, избежать множественного тестирования и сильно завышенных ошибок первого рода²³. Мы провели те же модели с различными уровнями сравнения (например, 4 против 2 опытных крыс и полные сиблинги против отсутствия сиблингов). Это не увеличивает частоту ошибок первого рода и не является множественным тестированием, поскольку мы не использовали разные модели с разным набором переменных. Мы оценили стабильность модели на основе DFBetas и сообщили минимальные и максимальные значения, которые представляют минимальные и максимальные значения разницы в каждой оценке параметра с каждой точкой данных и без нее. Мы использовали R версии 4.0.3⁶⁶ с функциями frailtyPenal из пакетов «frailtypack»⁶⁷,⁶⁸, «survival»⁶⁹,⁷⁰, «coxme»⁷¹, «survminer»⁷², «lme4»⁷³, «ggplot2»⁷⁴ и «multcomp»⁷⁵. На протяжении всей статьи средние значения и оцененные коэффициенты сообщаются с их стандартной ошибкой, если не указано иное; был принят уровень альфа 0,05.
Дополнительная информация
Данные
Мы загрузили коды (.r) и файлы данных (.RData) для подачи. В случае принятия они будут добавлены в общедоступный репозиторий или как часть дополнительных материалов. Коды можно открыть в R или Rstudio. Все файлы данных являются файлами .RData, которые можно открыть в R или Rstudio. Коды для открытия файлов данных находятся в файле кодов, например, load(“latency_first_success_naive_rats.RData”). Если у вас возникнут проблемы с открытием файлов данных, отправьте электронное письмо SCE.
Ссылки
Загрузка ссылок
Финансирование
Открытый доступ к финансированию был обеспечен и организован Проектом DEAL. Финансирование было предоставлено Швейцарским национальным научным фондом (номер гранта 31003A_176174) М.Т. Мы благодарим Фрейю Еву Николас Пуппо за помощь в сборе данных во время пилотного исследования. Мы благодарим Эви Цвиггарт за помощь в уходе за животными.
Информация об авторах
Авторы и их место работы:
- Кафедра социобиологии и антропологии, Институт зоологии и антропологии им. Иоганна Фридриха Блюменбаха, Геттингенский университет, 37077, Геттинген, Германия
- Саша С. Энгельхардт
- Кафедра этологии и социобиологии, Немецкий центр приматов, Лейбницкий институт приматологических исследований, 37077, Геттинген, Германия
- Саша С. Энгельхардт
- Поведенческая экология, Институт экологии и эволюции, Бернский университет, 3032, Хи́нтеркаппелен, Швейцария
- Саша С. Энгельхардт, Харшкумар Васойя и Михаэль Таборски
- Группа «Animal Minds», Институт нейронаук, Автономный университет Барселоны, Беллатерра, Барселона, 08193, Испания
- Харшкумар Васойя
- Кафедра коллективного поведения, Институт поведенческих наук им. Макса Планка, 78467, Констанц, Баден-Вюртемберг, Германия
- Михаэль Таборски
Авторы:
- Саша С. Энгельхардт
- Харшкумар Васойя
- Михаэль Таборски
Вклад:
С.С.Э. и М.Т. разработали исследование. С.С.Э. и Х.В. собрали данные из видеофайлов. С.С.Э. провела статистический анализ. Первоначальный черновик был написан С.С.Э., и все авторы пересмотрели рукопись. М.Т. руководил исследовательским проектом.
Корреспондирующий автор:
Sacha C. Engelhardt
Декларации об этике
Конфликт интересов
Авторы заявляют об отсутствии конфликта интересов.
Декларации об этике
Лицензия на проведение экспериментов с животными была выдана Швейцарским федеральным ветеринарным управлением кантона Берн (номер лицензии BE 55/18) М.Т. Разрешение на проведение необходимых исследований было выдано Бернским университетом (номер разрешения EAC201216T#212) М.Т.
Дополнительная информация
Издательское примечание: Springer Nature остается нейтральной в отношении юрисдикционных претензий в опубликованных картах и институциональных принадлежностях.
Дополнительные материалы
Ниже приведена ссылка на электронные дополнительные материалы.
Дополнительный материал 1
Дополнительный материал 2
Дополнительный материал 3
Дополнительный материал 4
Дополнительный материал 5
Дополнительный материал 6
Дополнительный материал 7
Дополнительный материал 8
Дополнительный материал 9
Дополнительный материал 10
Права и разрешения
Открытый доступ Этот документ лицензирован в соответствии с Международной лицензией Creative Commons Attribution 4.0, которая разрешает использование, обмен, адаптацию, распространение и воспроизведение в любом формате или на любом носителе, при условии, что вы соответствующим образом указываете авторство и источник, предоставляете ссылку на лицензию Creative Commons и указываете, были ли внесены изменения. Изображения или другие материалы третьих лиц, включенные в этот документ, относятся к лицензии Creative Commons, если иное не указано в подписи к материалу. Если материал не включен в лицензию Creative Commons статьи, и предполагаемое вами использование не разрешено законом или превышает допустимое использование, вам потребуется получить разрешение непосредственно от правообладателя. Чтобы просмотреть копию этой лицензии, посетите http://creativecommons.org/licenses/by/4.0/.
Перепечатка и разрешения
Об этой статье
Цитировать эту статью:
Engelhardt, S.C., Vasoya, H. & Taborsky, M. Experimental evidence for social learning in seminatural, wildtype Norway rats. Sci Rep 15, 37364 (2025). https://doi.org/10.1038/s41598025253166
Скачать цитату
Получено: 02 июля 2025 г.
Принято: 20 октября 2025 г.
Опубликовано: 27 октября 2025 г.
DOI: https://doi.org/10.1038/s41598025253166
Поделиться этой статьей
Любой, с кем вы поделитесь по следующей ссылке, сможет прочитать этот контент:
Получить ссылку для обмена
К сожалению, ссылка для обмена в настоящее время недоступна для этой статьи.
Копировать ссылку для обмена в буфер обмена.
Предоставлено инициативой Springer Nature по обмену контентом SharedIt.
Ключевые слова
Когнитивные способности, Приобретение информации, Инновации, Обучение, Социальная передача.