Интересное сегодня

Большая часть наших долгосрочных знаний организована в сложные сети. Считается, что сон играет критическую роль в абстрагировании знаний и усилении памяти на важные элементы для долгосрочного сохранения. Таким образом, сон должен способствовать развитию памяти на сети и абстрагированию их структуры для эффективного хранения. Однако это остается неизвестным, поскольку прошлые исследования сна фокусировались на дискретных элементах. Здесь мы исследовали влияние сна (парадигма внутрисубъектного исследования "ночной сон/дневное бодрствование" с участием 25 мужчин) на память о графовых сетях, где некоторые элементы были важны из-за плотных локальных связей (центральность по степени) или, независимо, важны из-за более сильных глобальных связей (центральность по близости/посредничеству). Сеть из 27 планет (узлов), слабо взаимосвязанных 36 телепортами (ребрами), была изучена посредством дискретных ассоциаций без явного указания какой-либо сетевой структуры. Несмотря на эквивалентное воздействие на все связи в сети, мы обнаружили, что память о связях между элементами с высокой локальной или высокой глобальной связностью лучше сохранялась после сна. Эти результаты подчеркивают, что сон обладает способностью усиливать как глобальную, так и локальную структуру из мира и абстрагировать множественные опыты для эффективного формирования внутренних сетей знаний.
Введение: Роль сна в консолидации знаний
Сон продемонстрировал свою способность поддерживать консолидацию декларативных воспоминаний. Ночь сна, как правило, улучшает память по сравнению с аналогичным периодом бодрствования днем. Основным механизмом, лежащим в основе этой консолидации, считается повторная реактивация недавно закодированных воспоминаний во время сна. Со временем реактивация перекрывающейся информации приводит к абстрагированию памяти, при этом некоторые детали теряются, а сохраняется суть опыта или центрально важная информация. Считается, что сон особенно важен для извлечения такой сути и построения схем. В соответствии с этим, было показано, что важные элементы, закодированные перед сном, больше усиливаются сном. Кроме того, сила памяти и сложность элемента влияют на то, насколько сон улучшает память, причем наибольшую пользу получают элементы с низкой силой и высокой сложностью.
Хотя большинство исследований сна фокусировались на дискретных элементах, таких как списки слов или пары элементов, большая часть информации в реальном мире взаимосвязана и интегрирована в сети знаний. Таким образом, остается неясным, как сон влияет на обучение сетевых структур. Недавно было высказано предположение, что гиппокамп и парагиппокампальные структуры, поддерживающие пространственную память и навигацию, могли эволюционировать у человека, чтобы в более широком смысле поддерживать обучение сетей знаний. Это распространилось и на концепции в обучении с подкреплением, где необходимо разрабатывать оптимальные стратегии для обучения новой информации.
Записи из отдельных клеток в гиппокампально-парагиппокампальной сети предоставили свидетельства того, что нейроны с определенной настройкой поддерживают аспекты представления когнитивной карты окружающей среды. Гиппокампальные клетки места у крыс демонстрируют пространственно локализованные паттерны активности во время движения по средам, причем каждая клетка места активна в различных конкретных регионах среды. Коллективно они предоставляют уникальный код для каждого встреченного в среде местоположения. Во время периодов сна и неподвижности субпопуляции клеток места имеют тенденцию реактивироваться, причем порядок активных клеток "повторяет" последовательность посещенных мест в среде ранее. Такая реактивация, по-видимому, сохраняет топологическую структуру среды, причем последовательности реактивации по маршрутам в Y-образной лабиринте соответствуют физическим связям внутри среды. Это предполагает, что во время сна гиппокампальные сети будут реактивировать различные пути, пройденные в состоянии бодрствования, сохраняя структуру, и могут реактивировать пересечения или пути, которые встречаются чаще, если реактивация связана с объемом воздействия. Например, многократное прохождение через центральный узел при исследовании сети путей приведет к тому, что реактивации будут проходить через этот узел намного чаще, чем другие регионы.
Используя кинематографическую симуляцию сложной сети недавно изученных городских улиц, стало возможным изучить вызываемые гиппокампальные реакции на уличные сети во время навигации. При входе в новые уличные перекрестки, если новая улица содержала больше локальных улиц на выбор (более высокая центральность по степени), активность заднего гиппокампа увеличивалась, а если вариантов было меньше (например, тупик), активность заднего гиппокампа снижалась. В то время как активность заднего гиппокампа реагировала на локальную связность, передний гиппокамп реагировал на изменения глобальной связности (центральность по близости). Его активность увеличивалась при входе в более глобально связанную улицу в сети и снижалась при входе в менее глобально связанные улицы. Кроме того, исследование, в котором участники обучались находить маршруты через небольшую сеть улиц в среде виртуальной реальности, показало, что повышенная активность гиппокампа во время сна связана с улучшением производительности после сна. Та же группа позже продемонстрировала, что сон может переструктурировать навигационное поведение, чтобы оно стало менее зависимым от стратегии, зависящей от пространственного гиппокампа, и задействовало дополнительную стратегию, основанную на реакции на стимулы, связанную со стриатумом. В совокупности эти исследования предполагают, что на следующий день после изучения уличной сети гиппокамп способен отслеживать связность в сети во время навигации, и что навигационные воспоминания реактивируются и трансформируются во время сна. Мы только начинаем понимать, как информационные сети и их топологическая структура изучаются и консолидируются в промежутке между обучением и навигацией.
Недавно ряд исследований начал исследовать, как могут быть изучены графовые структуры. Однако большинство этих исследований отслеживали память только в течение короткого периода времени или исследовали только обучение. В отличие от этого, недавнее исследование предложило участникам изучить структурированную информацию в соответствии с графом и извлечь ее через 24 часа в сканере МРТ. Нейронная активность, измеренная в энторинальной коре, отслеживала расстояние между элементами в изученном графе. Другое интересное исследование (однако с коротким интервалом сохранения, составляющим минуты) исследовало, как локальная связность, т.е. структура сообщества, влияет на статистическое обучение, и показало, что участники чувствительны к этому типу топологии, поскольку они идентифицировали ребра, соединяющие локальные сообщества, как естественные точки разрыва. Однако, насколько нам известно, не было проведено исследований влияния локальной и глобальной связности (т.е. центральности по степени и центральности по близости) на обработку информации и приобретение памяти в долгосрочной перспективе. Также не исследовалось, как сон может влиять на обучение сетей, где теории подчеркивают важность извлечения сути из опыта, что, по-видимому, связано со связностью узлов в сети.
Цели исследования
Здесь мы исследуем, как сон во время сохранения влияет на ассоциации, изученные с использованием задачи явного обучения графа с топологией, которая позволила нам разделить вклады локальной и глобальной связности. Мы ожидали, что:
- (1) Более слабые/более сложные ассоциации будут больше улучшаться сном во время интервала сохранения (как было продемонстрировано в других местах, т.е. большее расстояние между узлами будет прогнозировать большую пользу от сна во время интервала сохранения).
- (2) Важная информация будет больше улучшаться сном во время интервала сохранения, так что высокая центральность (локальная или глобальная связность) будет прогнозировать большую пользу от сна во время интервала сохранения.
В нашем дизайне ребра, связанные с узлами высокой и низкой центральности, систематически не отличались по воздействию во время обучения, поскольку мы тщательно и псевдослучайным образом выбирали маршруты. Поскольку релевантность, как было показано, обычно усиливает эффект сна, мы не ожидали никаких различий между узлами высокой глобальной или локальной связности в их восприимчивости к эффектам памяти, зависящим от сна, но что они будут одинаково усилены сном по сравнению с узлами низкой глобальной или локальной связности. Кроме того, мы противопоставили релевантность, полученную из центральности, более классической релевантности, связанной с подкреплением, связав некоторые узлы с денежным вознаграждением и наказанием. Чтобы предотвратить смешивание эффектов центральности и подкрепления, мы построили граф с тройной вращательной симметрией и разместили вознагражденные, наказанные и нейтральные узлы в симметричных положениях внутри графа.
Примечание: В этом первоначальном эксперименте мы включили только мужчин, чтобы повысить вероятность обнаружения предполагаемых различий между сном и бодрствованием. В будущих исследованиях мы планируем обобщить эти результаты на женщин.

Методы исследования
Участники
В исследовании приняли участие двадцать пять здоровых молодых мужчин в возрасте от 18 до 30 лет (средний возраст 24,20 ± 3,53 года). Женщины показали различия в обучении и памяти в целом и в консолидации, зависящей от сна, в частности, в течение менструального цикла, а высокая временная интенсивность исследований сна ограничивает возможность увеличения мощности путем увеличения размера выборки. Следовательно, мужчины были выбраны для этого исследования, чтобы уменьшить вариативность в выборке, что оптимизировало статистическую мощность в этих рамках. Примечательно, что простое добавление двадцати пяти женщин к нашей выборке не позволило бы нам надежно выявить различия между мужчинами и женщинами, поскольку это был бы эффект внутрисубъектного и междусубъектного взаимодействия, а также эффект более высокого порядка, который, скорее всего, потребовал бы гораздо больше участников для обнаружения. Участники не курили, свободно владели английским языком, не принимали в настоящее время лекарства и не имели никаких физических или психических расстройств. Все они сообщили о регулярном графике сна, ложась спать до полуночи (11:18 ± 47 мин) и просыпаясь до 8:00 утра (7:41 ± 47 мин). Кроме того, участники не работали в ночные смены, не имели диагностированных нарушений сна и не пересекали часовые пояса. Наконец, они не сообщали о каких-либо стрессовых событиях, таких как экзамены или дедлайны, до или во время эксперимента. Эксперимент был одобрен комитетом по этике UCL (номер ID: 8951/002), и все исследования проводились в соответствии с соответствующими руководящими принципами и правилами. Письменное информированное согласие было получено от каждого участника перед началом эксперимента. Участники были финансово вознаграждены за свое участие.
Дизайн и процедура
Исследование проводилось по сбалансированному внутрисубъектному дизайну, где участники проходили два сеанса, разделенных не менее чем 7 днями (8,44 ± 2,98 дня). Каждый сеанс состоял из фазы обучения и фазы извлечения с интервалом сохранения 10 часов между двумя фазами. В конце фазы обучения участникам было предложено избегать изучения новой информации или учебы и не повторять изученную информацию. В эксперименте для каждого участника тестировались два условия: сон и бодрствование. В условии бодрствования участники приходили в 8 утра, чтобы завершить фазу обучения, и возвращались в 8 вечера для фазы извлечения. В условии сна участники приходили в 8 вечера и возвращались на следующее утро в 8 утра. Последовательность условий была сбалансирована среди участников.
Эксперимент был разделен на две фазы. В первой, фазе обучения, участники завершили задачи обучения и вознаграждения, а во второй, фазе извлечения, они выполнили задачу вспоминания. В конце каждой фазы проводились контрольные измерения. Настроение участников оценивалось путем заполнения шкалы позитивного и негативного аффекта (PANAS), их субъективная сонливость измерялась по шкале сонливости Стэнфорда (SSS), а скорость реакции (1/время реакции, т.е. обратное время реакции) – объективная мера бдительности – была получена из 5-минутной версии психомоторного теста на бдительность (PVT). Мы выбрали 5-минутную версию вместо более длинных версий PVT, поскольку она снижает нагрузку на участников и обеспечивает достаточную мощность для обнаружения эффектов депривации сна, если скорость реакции (1/время реакции) используется как переменная исхода. Кроме того, в конце каждой фазы извлечения участники выполняли задачу генерации слов для оценки их способности извлекать высоко консолидированную информацию. Генерация слов является стандартной мерой, используемой во многих исследованиях сна и памяти, а также успешно использовалась для выявления долговременных дефицитов памяти при очень легких случаях деменции.
Наконец, в конце второй фазы извлечения участники заполнили шкалу Санта-Барбара SenseOfDirection (SBSOD), задавая вопросы о пространственных и навигационных способностях, и заполнили опросник навигационных стратегий (NSQ), задавая вопросы об их опыте навигации и их навигационной стратегии.
Структура графа
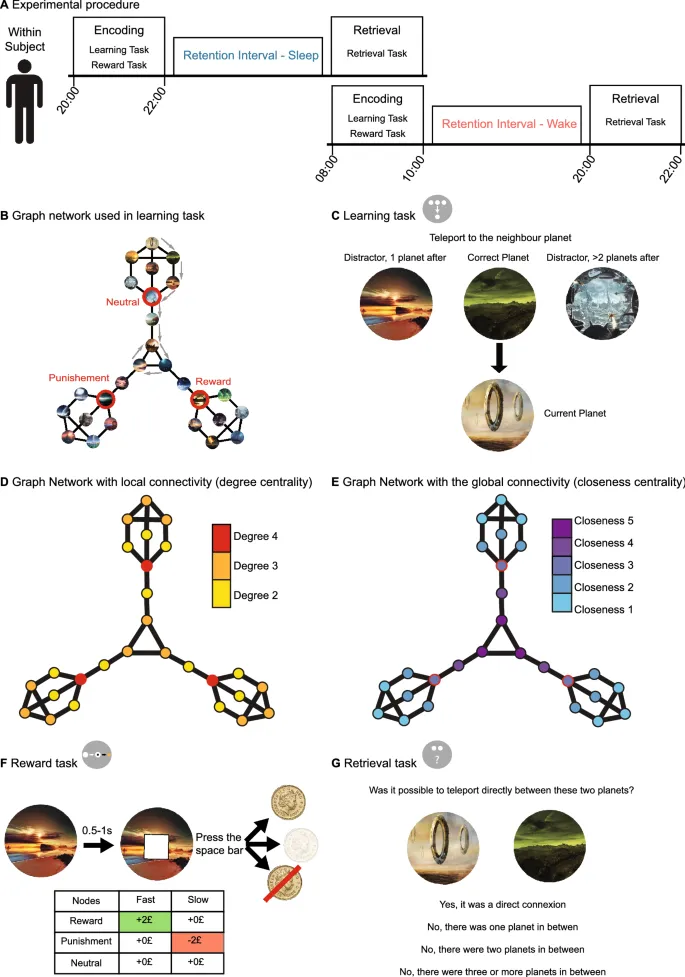
Был построен граф, состоящий из 27 узлов (представление графа см. на Рис. 1B), и каждому узлу были назначены изображения (уникальные ландшафты внеземных планет). Количество узлов было выбрано таким образом, чтобы обеспечить эффективное кодирование в течение 1,5 часов фазы обучения (по результатам пилотных участников). Граф содержал 36 ребер, соединяющих узлы. В процессе построения графовые параметры центральности по близости и центральности по степени были ортогонализованы, и преследовалась тройная симметрия. Кроме того, три узла, соответствующие красным узлам на Рис. 1B, были выбраны для усиления. Узлы были либо положительно усилены (узел вознаграждения), отрицательно усилены (узел наказания) или не усилены (нейтральный узел). Усиления ассоциировались во время задачи вознаграждения. Этот граф никогда не показывался участникам во время эксперимента, и участникам явно не сообщалось о базовой структуре задачи обучения.
Задача обучения
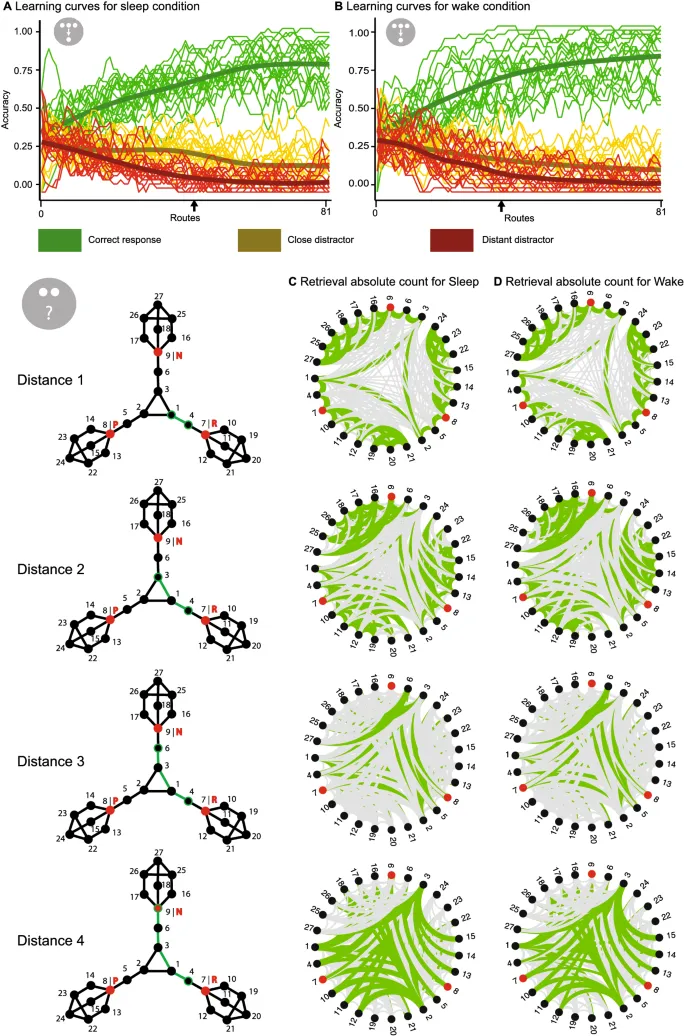
Задача обучения была геймифицирована для оптимизации мотивации участников во время обучения и извлечения. Кратко, задача была встроена в сюжет о человечестве на грани вымирания на Земле, и участники исследовали планеты, чтобы найти новый дом для жизни людей. Наше пилотирование продемонстрировало, что мотивация участников, особенно во время 1,5-часовой сессии обучения, значительно выиграла от этого подхода, что позволило нам использовать несколько больший граф. Чтобы ознакомить участника со стимулами, каждая планета отображалась в середине экрана с ее названием под ней в течение 2 секунд с интервалом между стимулами 0,5 секунды. Затем участники изучали структуру графа, выполняя 81 маршрут длиной 8 переходов между планетами графа. Обзор задачи можно увидеть на Рис. 1. Для каждого перехода участникам предлагалось идентифицировать соседа текущей планеты, представленного с его названием внизу экрана (т.е. планета, связанная одним ребром), при этом им показывалось три потенциальные планеты с их названиями наверху экрана. Один вариант был правильной планетой (одна из 2-4 связанных планет), а два других варианта были неправильными (т.е. не были непосредственно связаны). Из двух неправильных выборов один представлял планету на расстоянии двух ребер, т.е. между текущей планетой и неправильным выбором была одна планета, а другой – на расстоянии трех или более ребер, т.е. между ними было по крайней мере две планеты. Неправильные выборы выбирались случайным образом из всех планет, соответствующих аргументу расстояния, и учитывалось только кратчайшее расстояние для этого выбора. Во время каждого перехода у участников было максимум 10 секунд, чтобы выбрать правильный переход. Если они не отвечали в пределах временного лимита, выбор считался неправильным, и представлялся следующий испытание.
Задача вознаграждения
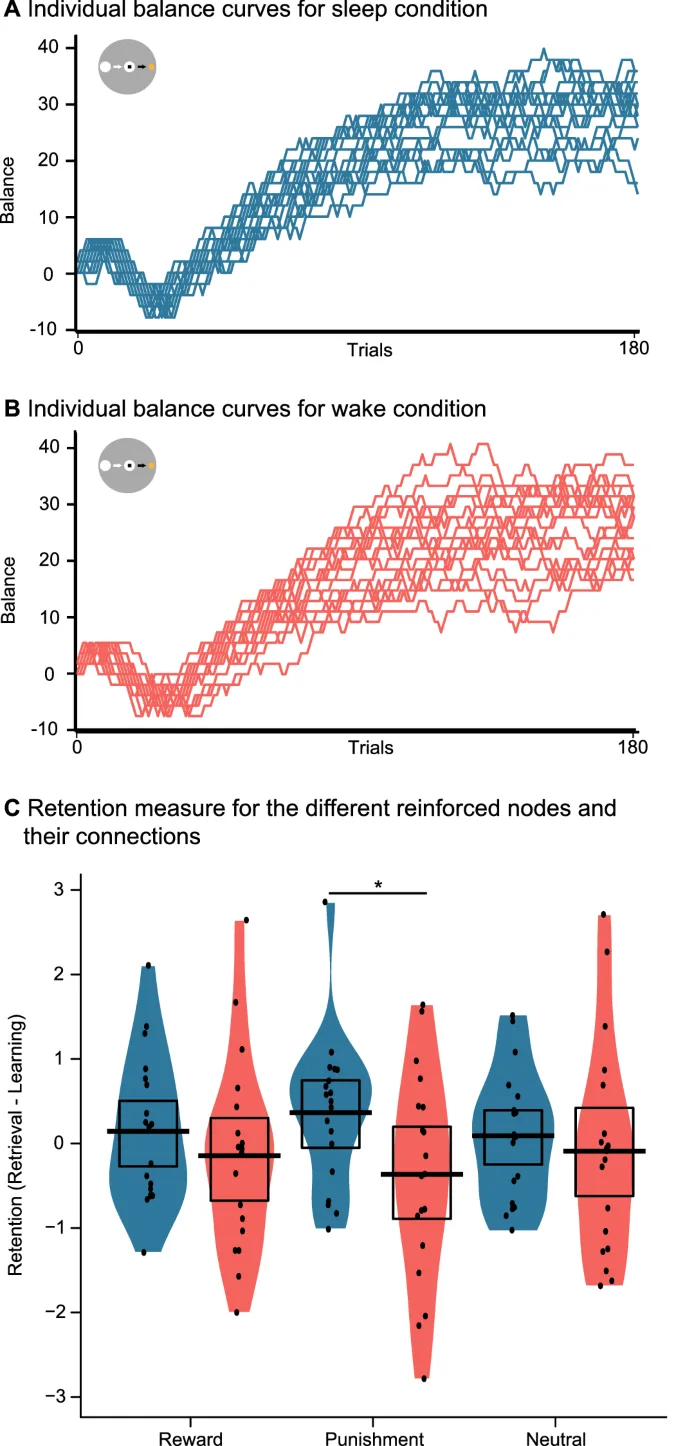
Задача вознаграждения была построена как адаптация задачи задержки денежного стимула, которая надежно активирует области вознаграждения. Во время задачи участники видели три упомянутых выше усиленных узла. В течение 180 испытаний фиксирующий крест появлялся в центре экрана в течение 250 мс, затем одна из трех планет, представляющих узлы, в течение 2 секунд. Через 0,5-1 секунду появлялся белый квадрат, и участникам нужно было как можно быстрее нажать пробел (нажатие до появления квадрата считалось промахом). В зависимости от их времени реакции (RT) и представленной планеты, участники получали различные денежные результаты. Для планеты вознаграждения, если участники были достаточно быстры, они получали +2 фунта стерлингов, а если недостаточно быстры – 0 фунтов стерлингов. Для планеты наказания, быть достаточно быстрым означало заработать 0 фунтов стерлингов, а быть слишком медленным – потерять −2 фунта стерлингов. Наконец, для нейтральной планеты участники получали 0 фунтов стерлингов независимо от того, были они быстры или нет. Их выигрыши и проигрыши накапливались, напоминая сумму, которую они получили бы за выполнение задачи вознаграждения.
Задача извлечения
Задача извлечения была разделена на три части. В первой части мы представляли две планеты (без их названий) бок о бок и просили участников решить, были ли планеты напрямую связаны во время обучения, или между ними была одна, две, три или более планет. Это делалось для всех возможных комбинаций 27 планет, поэтому участникам было представлено 351 испытание (если бы участники выбирали совершенно случайным образом, они были бы правы в 25% случаев). Во второй части мы спрашивали участников о названиях планет. Участники выполнили 27 испытаний, по одному для каждой планеты, и их спрашивали, какое из четырех возможных названий было правильным. Три неправильных названия были выбраны из планет, находящихся на расстоянии одного, двух или более ребер соответственно. В последней части участник определял условные зависимости, изученные в задаче вознаграждения. Это означает, что им показывали различные планеты, использованные для задачи вознаграждения, и просили определить нейтральную, вознаграждающую и наказанную планету соответственно.
Обработка данных и статистический анализ
Данные 6 участников были исключены из анализа. Четыре участника имели производительность обучения с точностью ниже 0,5, а два участника имели высокую производительность обучения, точность выше 0,8, но низкую производительность извлечения, менее 0,4. Мы исключили участников с низким уровнем обучения (т.е. не существенно выше случайного уровня 33%), поскольку они имели бы мало или совсем не знали о базовой структуре графовой сети, что было бы необходимо для воздействия топологии узлов. Мы исключили участников с высоким уровнем обучения, но низкими показателями извлечения, поскольку эти участники, вероятно, не усвоили ассоциации более высокого порядка, необходимые для высокой производительности при извлечении, и, следовательно, также не смогли бы полагаться на знания о графе. Эти запланированные исключения произошли до сравнения показателя сохранения, нашей основной переменной исхода, между сном и бодрствованием. Кроме того, анализировались только пары узлов с расстоянием от 1 до 4, поскольку извлечение проверяло знание участниками структуры графа только до расстояния 4 максимум. Обработка данных проводилась в Matlab 2018a, а статистический анализ зависел от R studio (версия 1.0.143). Анализ основывался в основном на дисперсионном анализе с повторными измерениями, парных t-тестах, корреляциях Пирсона и регрессии с линейным моделированием. Подробности обработки данных и статистического анализа можно найти в дополнительных методах.
Результаты
Производительность сохранения
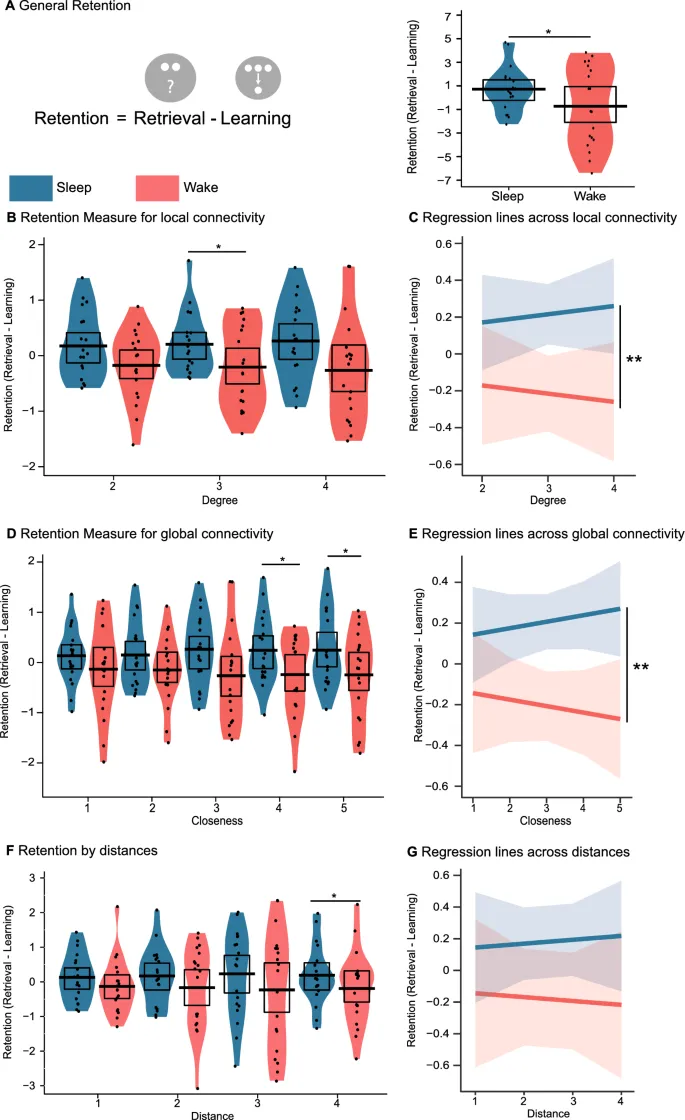
Показатель сохранения был создан путем вычитания производительности обучения из результатов извлечения, т.е. сколько информации участник сохранил в течение периода сохранения. Важно отметить, что производительность обучения и производительность извлечения были высоко коррелированы (r = 0,88 и r = 0,87 для сна и бодрствования соответственно), что указывает на то, что показатель сохранения является действительной мерой исхода. При усреднении всей информации о расстоянии мы обнаружили, что участники сохранили больше информации от обучения до извлечения в условиях сна, чем в условиях бодрствования (t(18) = 2,13; p = 0,048). Дисперсионный анализ по расстояниям подтвердил этот основной эффект интервала (F(1,18) = 4,52; p = 0,048). При индивидуальном анализе расстояний мы обнаружили, что для расстояния 4 в условиях сна также наблюдалось лучшее сохранение (t(18) = 2,29; p = 0,035), однако такое различие не было обнаружено для расстояний 1 (t(18) = 1,01; p = 0,327), 2 (t(18) = 1,33; p = 0,201) и 3 (t(18) = 1,37; p = 0,188).
Используя линейное регрессионное моделирование, которое в отличие от дисперсионного анализа позволяло нам учитывать порядок предиктора расстояния, расстояния по интервалу выявили различие в интерцепте между условиями сна и бодрствования, которое можно интерпретировать так же, как основной эффект в дисперсионном анализе (F(1,18) = −3,99; p = 0,016). Однако этот анализ не выявил различий в наклонах, что можно интерпретировать так же, как взаимодействие расстояния и интервала в дисперсионном анализе (F(1,18) = −2,22; p = 0,091). Важно отметить, что, поскольку ни дисперсионный анализ, ни регрессионные модели не обнаружили свидетельств взаимодействия расстояния и интервала, следует интерпретировать только основной эффект сна, а пост-хок парные сравнения включены только для полноты.
В отношении локальной связности (центральность по степени) участники сохранили больше информации во время интервала сна (F(1,18) = 4,71; p = 0,044). Этот эффект был сильнее для узлов с центральностью по степени 3 (t(18) = 2,15; p = 0,046) по сравнению с центральностью 2 (t(18) = 2,01; p = 0,060) или 4 (t(18) = 1,89; p = 0,075). Регрессионный анализ подтвердил основной эффект сна (разница в интерцепте: F(1,18) = −4,86; p = 0,039) и указал, что более высокая центральность по степени была связана с увеличением пользы от сна во время сохранения (разница в наклоне: F(1,18) = −8,26; p = 0,015).
Аналогичные результаты были получены для глобальной связности (центральность по близости), поскольку участники снова показали лучшие результаты в течение интервала сохранения сна (F(1,18) = 5,26; p = 0,035), что отразилось в пользе сна для центральности по близости 4 (t(18) = 2,27; p = 0,036) и 5 (t(18) = 2,90; p = 0,001), но не для центральности по близости 1 (t(18) = 1,10; p = 0,285), 2 (t(18) = 1,41; p = 0,174) или 3 (t(18) = 1,89; p = 0,075). Опять же, регрессионный анализ показал, что участники показали лучшие результаты во время сна (разница в интерцептах: F(1, 18) = −3,79; p = 0,009), и что более высокая центральность по степени увеличила эффект сна (разница в наклонах: F(1, 18) = −3,59; p = 0,012).
Производительность обучения и извлечения
Что касается результатов обучения, то описательно общая производительность обучения (рассчитанная путем усреднения результатов кодирования по четырем расстояниям) была выше в условиях бодрствования, чем во сне, но эта разница не была статистически значимой (t(18) = −2,09; p = 0,051). Однако, рассматривая только связи первого порядка (как они были изучены во время задачи) и разделяя задачу обучения на три части, не было выявлено основного эффекта сна или бодрствования при анализе трех частей в дисперсионном анализе (F(1,18) = 2,66; p = 0,120) или для последней трети в индивидуальном t-тесте (t(18) = −1,09; p = 0,289). Хотя участники увеличивали свою производительность обучения по частям (F(1, 18) = 124,74; p < 0,001). Индивидуальные кривые обучения участников показаны на Рис. 3A,B для обоих условий.
Что касается локальной связности, дисперсионный анализ показал, что более высокая центральность по степени была связана с лучшим обучением (F(1, 18) = 44,55; p < 0,001), но не было эффекта интервалов (F(1, 18) = 4,17; p = 0,056) или взаимодействия (F(1, 18) = 0,63; p = 0,540). Для глобальной связности были получены аналогичные результаты, т.е. участники лучше изучали узлы с более высокой центральностью по близости (F(1, 18) = 33,01; p < 0,001), но не было эффекта для интервала сохранения (F(1, 18) = 4,33; p = 0,052) и не было взаимодействия (F(1, 18) = 1,56; p = 0,195). Аналогично, не было выявлено статистических различий для анализа линейной модели. Что касается общей производительности извлечения (рассчитанной по частоте правильных ответов для каждого расстояния), участники были лучше в правильном определении близких пар, чем более отдаленных (F(1,18) = 7,46; p < 0,001), но не было основного эффекта интервала (F(1,18) = 0,37; p = 0,551) или эффекта взаимодействия (F(1,18) = 0,13; p = 0,939). Визуализация необработанных данных извлечения также может быть найдена на Рис. 3C,D. Не было найдено статистических различий для локальной связности (F(1, 18) = 2,88; p = 0,069), но глобальная связность показала основной эффект центральности (F(1, 18) = 4,34; p = 0,003). Аналогично обучению, статистических различий для анализа линейной модели не было. Наконец, участники не смогли лучше запомнить ассоциации названий планет в условиях сна, чем бодрствования (t(18) = 0,89; p = 0,384).
Подкрепление
Индивидуальные кривые баланса (сумма денег, выплаченная участникам) для двух условий можно найти на Рис. 4A,B. Что касается показателя сохранения, дисперсионный анализ не выявил влияния сна или бодрствования (F(1, 18) = 2,33; p = 0,144), а также не было эффекта подкрепления (F(1,18) = 0; p = 1) или взаимодействия двух (F(1,18) = 1,12; p = 0,337). Однако разведочные парные t-тесты показали, что для узла наказания участники показали лучшие результаты в условиях сна (t(18) = 2,10; p = 0,049), но не для вознаграждения (t(18) = 0,92; p = 0,368) и нейтральных узлов (t(18) = 0,49; p = 0,629). Обратите внимание, что участники могли явно вспомнить условные зависимости вознаграждения после задачи подкрепления, что было проверено следователем путем демонстрации трех планет и вопроса о связанных денежных исходах.
Навигационные тесты
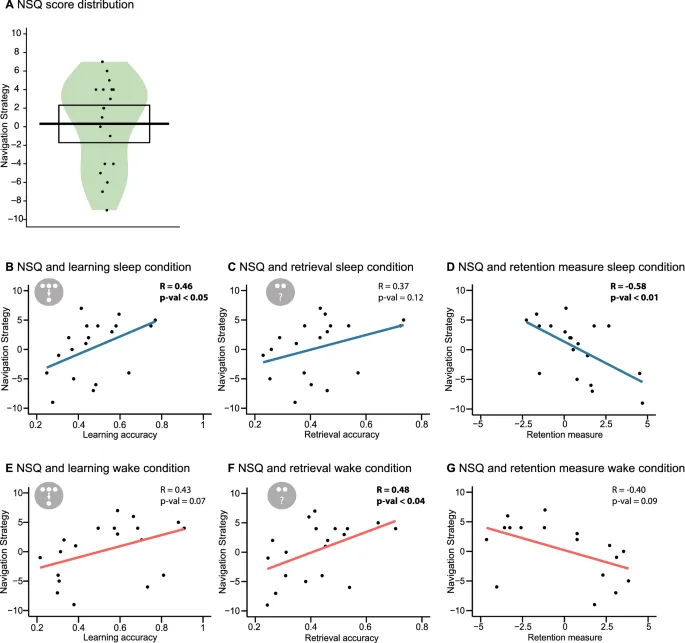
Была найдена значимая корреляция между двумя навигационными опросниками (r = 0,55, p = 0,015). Поскольку вариативность NSQ была больше, дальнейшие корреляции использовали этот навигационный тест. Участники с более высоким показателем по NSQ имеют более высокую картографическую стратегию. Мы обнаружили значительную отрицательную связь между NSQ и показателем сохранения для сна (r = −0,58, p = 0,009), но не для условия бодрствования (r = −0,40, p = 0,093). Статистически значимых различий при сравнении коэффициентов корреляции с использованием процедуры Z Фишера не было (Z = −0,85, p = 0,393). Мы обнаружили положительную связь между NSQ и производительностью обучения в условиях сна (r = 0,46, p = 0,048) и между NSQ и производительностью извлечения в условиях бодрствования (r = 0,48, p = 0,036). Корреляции для производительности обучения в условиях бодрствования (r = 0,43, p = 0,066) и производительности извлечения в условиях сна (r = 0,37, p = 0,122) не достигли значимости. В целом, хотя некоторые связи не достигли значимости в условиях сна или бодрствования, общая картина эффектов была схожей. Кроме того, не было обнаружено различий между коэффициентами корреляции при сравнении корреляций обучения (Z = 0,23, p = 0,821) или извлечения (Z = −0,79, p = 0,432) между сном и бодрствованием.
Контрольные задачи
Не было никакой разницы в долгосрочной производительности извлечения (измеренной с помощью задачи генерации слов) между условиями сна и бодрствования (t(18) = −0,82; p = 0,423). Также не было обнаружено статистических различий для объективной бдительности (скорость реакции, т.е. обратное время реакции, PVT), субъективной сонливости (измеренной SSS) или позитивного или негативного аффекта (измеренного PANAS) между условиями сна и бодрствования во время обучения (PVT: t(18) = 1,02, p = 0,321; SSS: t(18) = −0,57, p = 0,578; PANASпозитивный: t(18) = 1,37; p = 0,188; PANASнегативный: t(18) = 2,08; p = 0,052) или извлечения (PVT: t(18) = 0,66, p = 0,519; SSS: t(18) = −0,86; p = 0,399; PANASпозитивный: t(18) = 0,67; p = 0,509; PANASнегативный: t(18) = −0,33; p = 0,742). Описательная статистика может быть найдена в Таблице 1.
Обсуждение
Здесь мы исследовали влияние сна на консолидацию изученных топологических сетей, которые различались по глобальной и локальной связности узлов. Мы обнаружили, что как связи с глобально, так и локально высокорелевантными узлами были предпочтительно усилены сном. Это произошло несмотря на равное воздействие на все связи в сети во время обучения. Напротив, сон не оказал никакого влияния на усиление узлов, сделанных заметными денежным подкреплением. Мы обсуждаем, как эти результаты помогают продвинуть наше понимание того, как репрезентации изученных графовых сетей затрагиваются оффлайн-обработкой, моделями сна и консолидации, а также их значением для понимания оффлайн-реактивации в гиппокампальных сетях.
Текущее исследование представляет собой новую задачу ассоциативного обучения, в которой информация изучалась в соответствии с графовой сетью. Мы обнаружили, что во время обучения граф влиял на поведение за пределами ассоциаций первого порядка, поскольку близкие дистракторы (дистракторы, находящиеся всего в одном ребре от прямой связи) чаще выбирались, чем далекие дистракторы (дистракторы, находящиеся на расстоянии не менее двух ребер от прямой связи) при ошибках участников. Хотя сон улучшил саму консолидацию памяти, его эффект не был усилен увеличенным расстоянием между узлами, как можно было ожидать, учитывая, что сон предпочтительно усиливает элементы с низкой ассоциацией и высокой сложностью. Однако сон специфически усилил элементы, которые были более релевантны для навигации по сети, поскольку узлы с высокой локальной и глобальной связностью показали более сильный эффект сна. Этот градиентный эффект топологической релевантности может быть связан с результатами градиентных эффектов вознаграждения на память в лабиринте. В этом исследовании участники исследовали лабиринт, открывая карточки, разложенные в 2D-сетке, и получали высокое или низкое вознаграждение после определенного количества карточек. Эффект вознаграждения был выше, чем ближе карточка была к финальной открытой карточке. Хотя мы обнаружили аналогичный градиентный эффект топологической релевантности, мы не обнаружили эффекта денежного подкрепления, примененного к подмножеству узлов. Это также может быть объяснено распространением вознаграждения по сети, если предположить, что наша сеть была слишком мала. Мы выбрали размер нашей сети после обширного пилотирования, чтобы ее можно было выучить примерно до 80% за 1,5 часа. Использование большей сети или, возможно, даже двух сетей с различными процедурами подкрепления может оказаться более плодотворным.
Предполагается, что сон усиливает абстракцию сути из эпизодов. Такая абстракция сути может стать более важной при изучении больших сетей, поскольку участники будут испытывать трудности с удержанием всех связей в памяти. Представление сети на разных масштабах тогда позволит более эффективную обработку памяти. Интеграция очень больших сетей может происходить в течение нескольких ночей сна, как было показано для других процессов абстракции сути. После построения большой сети может оказаться, что новые элементы могут быть добавлены к этой схеме гораздо быстрее и с меньшей зависимостью от гиппокампа. Фактически, сон может больше не требоваться для консолидации таких воспоминаний, поскольку кодирование может обойти гиппокамп, напрямую интегрируясь в неокортикальную сеть знаний. Однако, основываясь на предыдущих работах, демонстрирующих такое быстрое изучение новых узлов в сетях, мы бы предсказали, что после добавления новых узлов к нашей сети эта вновь закодированная информация потребует меньшей консолидации во сне для стабилизации, при условии, что глобальная топология минимально затронута, и, следовательно, системная консолидация будет завершена гораздо быстрее. Это можно было бы изучить в нашей парадигме, добавив узлы, которые сильно влияют или не влияют на топологию. Например, обеспечение одного ярлыка между очень далекими частями сети может радикально изменить глобальную связность узлов и потенциально привести к более продолжительной консолидации во время сна.
Одно из предсказаний теорий, подчеркивающих важность глобальной экстракции сути и развития схем, состоит в том, что преимущественно глобально важная информация будет приоритетной по сравнению с локально важной информацией. Мы не обнаружили, что это так, поскольку высокая локальная связность также повлияла на консолидацию во время сна. В будущих исследованиях было бы интересно изучить память через несколько дней, чтобы наблюдать, теряется ли глобальная и локальная информация или сохраняется с той же скоростью. Тестирование других сетевых структур также помогло бы исследовать, являются ли локальные и глобальные эффекты аддитивными, т.е. узлы с высокой локальной и глобальной центральностью удваиваются ли сном?
Согласно одной точке зрения, гиппокампальная реактивация во время сна считается тесно связанной с предыдущим опытом, так что последовательности клеток места, наблюдаемые во время обучения, снова появятся во время последующего сна. Было показано, что вознаграждение влияет на реактивацию клеток места, а увеличение вознаграждения увеличивает частоту событий реактивации в короткие периоды отдыха во время обучения. Другое исследование показало, что усиление дофаминергической модуляции в гиппокампе увеличивает частоту реактивации во время отдыха после кодирования. Альтернативная точка зрения недавно предположила, что в отсутствие вознаграждения события реактивации могут представлять случайные выборки из доступных траекторий через пространство. Усиление глобально релевантных узлов в нашей сети может быть объяснено любой из этих точек зрения. Либо глобальная релевантность была выведена уже во время бодрствования и усилена реактивация этих узлов во время сна механизмом синаптического тегирования, либо структура сети могла сместить случайные блуждания, происходящие в виде случайных блужданий по графу, чтобы подчеркнуть узлы с высокой центральностью по посредничеству. Поскольку центральность по посредничеству и центральность по близости были высоко коррелированы в нашем графе, мы не можем в настоящее время различить, какая из двух метрик может повлиять на консолидацию во время сна. С более крупной сетью можно было бы разделить центральность по близости и центральность по посредничеству. Если реактивация совершает случайные блуждания по сети, она может специально усиливать области с высокой центральностью по посредничеству, но не центральность по близости. Однако, возможно, что реактивация отдает приоритет важным структурам для изучения, что подтверждается недавними данными.
Здесь мы нашли предварительные данные (небольшая выборка корреляции), что участники, которые демонстрируют эффективное обучение графа, как правило, имеют более высокую самоотнесенную оценочную склонность к навигации с помощью карт и мышлению в терминах карт для навигации, основываясь на NSQ. Это перекликается с недавними данными о том, что люди, которые делают лучшие выводы о структуре графовых сетей, демонстрируют больше модельно-ориентированного планирования в многошаговом задании планирования. Будущие исследования с крупномасштабными онлайн-когортами были бы полезным способом исследования надежности таких корреляций и того, какие другие модерирующие факторы могут влиять на эти отношения. В этом отношении также было бы важно понять, представляет ли отрицательная связь между показателями NSQ и производительностью сохранения истинную связь, или же она обусловлена положительной связью как производительности обучения, так и извлечения.
Существует несколько ограничений текущей работы, которые следует учитывать. Мы выбрали сравнение дневного бодрствования и ночного сна, что несет в себе недостаток конфаунда циркадных ритмов. Выбор депривации сна против нормального сна вместо этого, как утверждается, вызывает проблему неспецифических эффектов на познание при извлечении. Это можно смягчить, добавив восстановительный сон, который, однако, показал снижение полезного эффекта сна, предположительно, позволяя отложенную консолидацию, зависящую от сна. Мы взвесили эти возможности при принятии решения, и статистически не было никакой разницы в производительности между условиями, где обучение происходило утром против вечера. Однако описательно производительность обучения была выше утром, и мы не можем исключить, что может существовать циркадный эффект на обучение, который мы не обнаружили из-за ограничений размера выборки. Хотя наш анализ основывался на разностных показателях, которые учитывают потенциальные различия в исходном уровне, остается возможность эффектов потолка в условии бодрствования. Мы также использовали процедуру извлечения, отличную от обучения. Мы сделали это, поскольку во время извлечения нас интересовали ассоциации более высокого порядка, выходящие за рамки ассоциаций первого порядка, которые были изучены. Добавление немедленного извлечения после обучения спутало бы эксперимент, поскольку тогда было бы неясно, действовала ли консолидация, зависящая от сна, на изученную информацию или на информацию, немедленно извлеченную. Хотя мы предприняли большие усилия при выборе топологии графа для этого эксперимента, чтобы ортогонализовать интересующие нас меры центральности (центральность по близости и центральность по степени), тестирование различных топологий графа выходило за рамки данного исследования, что поэтому ограничивает обобщаемость наших выводов. Кроме того, хотя тройная симметрия нашего графа использовалась для ограничения конфаунда усиленных узлов на результаты, связанные с центральностью, мы не можем полностью исключить, что наши результаты будут отличаться для графов, не имеющих усиленных узлов. Удивительно, но мы обнаружили, что производительность сохранения во второй экспериментальной сессии снизилась, а не увеличилась, как обычно ожидается при повторении задачи на память. Спекулятивное, но разумное объяснение этого может заключаться в том, что абстрагированная информация графа из первой сессии помешала сохранению структуры графа во второй экспериментальной сессии. Поскольку предполагается, что интерференция взаимодействует с эффектом сна, было бы интересно систематически исследовать этот эффект в будущем. Альтернативно, хотя мы постарались явно не информировать участников о структуре графа, процедура извлечения, вероятно, позволила им догадаться, что существует структура, выходящая за рамки парных ассоциатов, которая могла быть использована для стратегического улучшения памяти во время второй экспериментальной сессии. Однако, поскольку производительность сохранения снизилась во второй экспериментальной сессии, это кажется маловероятным. Мы также не обнаружили, что ассоциации названий планет были затронуты, что ожидалось бы, учитывая широкую поддержку памяти пар слов и других ассоциативных воспоминаний, которые выигрывают от сна во время интервала сохранения. Это могло быть связано с произвольностью имен или с тем, что другие компоненты задачи были более релевантны. Наш выбор задавать вопросы о планетах на расстоянии четырех или более ребер, но анализировать только до четырех ребер, создал ненужное бремя для участников и мог привести к предвзятости, поскольку большинство ответов были бы 4 или более. Будущие исследования должны собирать данные только для узлов на расстоянии до четырех шагов. Большие будущие исследования также могли бы пролить свет на различие между людьми, которые могли и не могли изучить задачу (4 участника не смогли учиться), а также теми людьми, которые могли изучить задачу, но не смогли извлечь многошаговые ассоциации более высокого порядка (2 участника).
В заключение, мы обнаружили, что локальные и глобальные аспекты связей между отдельными элементами задачи декларативной ассоциативной памяти определяют доступ к консолидации памяти, зависящей от сна. Этот подход имеет потенциал для более детального изучения того, как реактивация влияет на структуру знаний декларативной памяти.
Ссылки
(Список ссылок не предоставлен в исходном тексте)
Благодарности
Эта работа была поддержана грантами Немецкого исследовательского фонда (DFG; FE 1617/11 и FE 1617/21) для GBF и грантом James McDonnell Scholar Award для HJS. Мы хотели бы поблагодарить Тим Беренс и Мону Гарверт за их вклад в эту работу.
Финансирование
Открытый доступ финансируется и организуется проектом DEAL.
Информация об авторах
Авторы
- G. B. Feld
- M. Bernard
- A. B. Rawson
- H. J. Spiers
Вклад авторов
G.B.F. и H.J.S. разработали исследование. G.B.F. подготовил все материалы исследования. M.B. собрал данные. M.B. провел анализ под руководством G.B.F. M.B. подготовил все рисунки. A.R. поддержал анализ. G.B.F. и M.B. написали первый вариант рукописи. H.J.S. и A.B.R. критически пересмотрели рукопись. Все авторы рассмотрели рукопись.
Соответствующие авторы
Корреспонденция с G. B. Feld или H. J. Spiers.
Декларации об этике
Конкурирующие интересы
Авторы заявляют об отсутствии конкурирующих интересов.
Дополнительная информация
Примечание издателя Springer Nature остается нейтральной в отношении территориальных претензий, опубликованных на картах, и институциональной принадлежности.
Дополнительная информация
Дополнительная информация.
Права и разрешения
Открытый доступ Эта статья лицензирована в соответствии с Международной лицензией Creative Commons 4.0, которая разрешает использование, обмен, адаптацию, распространение и воспроизведение в любом носителе или формате, при условии, что вы должным образом указываете авторство и источник, предоставляете ссылку на лицензию Creative Commons и указываете, были ли внесены изменения. Изображения или другие материалы третьих сторон, включенные в эту статью, включены в лицензию Creative Commons статьи, если иное не указано в подписи к материалу. Если материал не включен в лицензию Creative Commons статьи, и ваше предполагаемое использование не разрешено законодательством или превышает разрешенное использование, вам потребуется получить разрешение напрямую от правообладателя. Чтобы просмотреть копию этой лицензии, посетите http://creativecommons.org/licenses/by/4.0/.
Перепечатки и разрешения
Об этой статье
Feld, G.B., Bernard, M., Rawson, A.B. et al. Sleep targets highly connected global and local nodes to aid consolidation of learned graph networks. Sci Rep 12, 15086 (2022). https://doi.org/10.1038/s41598022177472
Скачать цитату
Получено: 03 ноября 2021 г.
Принято: 30 июля 2022 г.
Опубликовано: 05 сентября 2022 г.
Версия записи: 05 сентября 2022 г.
DOI: https://doi.org/10.1038/s41598022177472
Поделиться этой статьей
Любой, кого вы поделитесь по следующей ссылке, сможет прочитать этот контент:
Получить ссылку для общего доступа
Извините, ссылка для общего доступа в настоящее время недоступна для этой статьи.
Копировать ссылку для общего доступа в буфер обмена
Предоставлено инициативой Springer Nature SharedIt по обмену контентом