
Интересное сегодня
Исследование эффективной связности ЭЭГ при визуально-моторной образности с использованием многошкальной символьной энтропии переноса
Введение
Визуально-моторная образность (ВМО) является важным компонентом моторной образности, обладающим потенциалом применения в интерфейсах мозг-компьютер (ИМК) и моторной реабилитации благодаря более низкой стоимости обучения по сравнению с кинестетической моторной образностью (КМО). Однако нейронные механизмы, лежащие в основе ВМО, особенно эффекты образности руки и перспективы (первая-лицо, 1pp, против третьего-лица, 3pp), остаются неясными. Данное исследование изучает эффективную связность ВМО ЭЭГ с использованием многошкальной символьной энтропии переноса. Анализ времени-частоты выявил выраженную событийно-связанную синхронизацию (ERS) в альфа- и высокобета-диапазонах, в то время как анализ связности подчеркнул сильный поток информации в теменно-затылочной сети. Примечательно, что доминирующие потоки информации, связанные с эффектом руки, были обнаружены между моторными и задними теменно-затылочными областями, в то время как эффект перспективы показал более заметное влияние. Образность 1pp значительно усилила нисходящую модуляцию затылочной коры, тогда как образность 3pp задействовала правую заднюю теменную область, предполагая более сильную обработку пространственной локализации. Эти результаты дают новое представление о различных нейронных механизмах ВМО и ее потенциальных применениях в когнитивной нейронауке и инженерии мозг-машина.
Моторная образность (МО) стала одной из ведущих парадигм в области интерфейсов мозг-компьютер (ИМК) благодаря своей естественной логике использования и широкому спектру потенциальных применений. МО включает в себя мысленное моделирование действия без фактического выполнения двигательного выхода. Её можно разделить на два основных типа: кинестетическая моторная образность (КМО), при которой индивиды представляют ощущения мышечных сокращений во время выполнения конкретного действия, и визуально-моторная образность (ВМО), при которой они визуализируют действие, как если бы наблюдали соответствующее изображение в своем сознании. Кроме того, ВМО подразделяется в зависимости от перспективы воображаемого действия: ВМО от первого лица (VMI1), где участники представляют действие со своей собственной точки зрения, и ВМО от третьего лица (VMI3), где они наблюдают за своими действиями с внешней точки, как будто смотрят на поведение другого человека.
За последние десятилетия исследования нейровизуализации выявили различия в активации мозга между различными модальностями МО, особенно в отношении КМО. Было показано, что КМО в основном задействует такие области, как дополнительная моторная область (SMA), дорсальная и вентральная премоторная кора (PMC), первичная моторная кора (M1), верхние и нижние теменные дольки, а также сенсомоторные области. В противоположность этому, для ВМО сообщалось о вовлечении областей, менее непосредственно связанных с движением, включая прецентральную извилину, заднюю теменную долю и затылочные области, а также различные подрегионы теменной доли в зависимости от перспективы. Тем не менее, несоответствия в экспериментальных протоколах и интерпретациях привели к тому, что КМО стала основным фокусом для изучения активности мозга, связанной с МО, и разработки последующих приложений, таких как системы ИМК на основе МО. Общепринято мнение, что КМО напоминает фактическое выполнение двигательной активности, сосредоточенной в областях моторной коры фронто-теменной сети, в то время как ВМО имеет больше сходств с наблюдением за движением, что подтверждается более сильным вовлечением задних теменно-затылочных областей.
Благодаря своей способности захватывать сигналы с высоким временным разрешением, неинвазивная электроэнцефалография (ЭЭГ) позволяет детально исследовать характеристики времени-частоты, связанные с МО. Широко сообщаемые пре-моторные событийно-связанные десинхронизации (ERD) и пост-моторные событийно-связанные синхронизации (ERS) в контралатеральной моторной коре во время КМО считаются одними из наиболее отличительных особенностей МО. Эти характеристики также широко применялись в исследованиях моторной реабилитации и интерфейсов человек-машина. Однако, демонстрирует ли ВМО схожие характеристики ЭЭГ, до сих пор является предметом дискуссий. В последние годы анализы функциональной связности приобрели известность благодаря выявлению топологических взаимосвязей между областями мозга, а методы направленной связности, предоставляющие информацию о направлении потока информации, использовались для исследования взаимодействий между областями мозга во время МО. Для КМО были идентифицированы как однонаправленная, так и двунаправленная связность во фронто-теменных областях, включая сильные связи от дорсальной премоторной коры (dPMA) к M1, от SMA к M1 и от вентральной премоторной коры (vPMA) к билатеральным верхним и нижним теменным долям. Тем не менее, эффективная сетевая динамика ВМО остается неясной, что затрудняет характеристику точной природы ВМО и ее представления в активности мозга.
По мере расширения исследований МО в области ИМК, более глубокое изучение ее физиологических и психологических механизмов несколько отставало. Один из ключевых вопросов заключается в том, может ли ВМО, учитывая ее гибкость и низкую стоимость, заменить КМО в практических контекстах, или она лучше подходит для приложений, использующих ее уникальные нейронные характеристики. Прежде чем ответить на эти вопросы, необходимо прояснить электрофизиологические основы ВМО.
С одной стороны, предыдущие исследования моторной образности преимущественно фокусировались на моторной коре и связанных с ней областях, часто рассматривая двигательное исполнение как базовый уровень для МО. Хотя этот метод применим к КМО, он может быть менее подходящим для исследования ВМО. Предыдущие исследования предполагали, что возбудимость моторной коры связана с КМО, но не обязательно с ВМО, указывая на то, что оценка ВМО через сенсомоторные ритмы может давать ограниченные или неубедительные результаты. Следовательно, данное исследование фокусируется на теменно-затылочных областях, часто упоминаемых в предыдущих отчетах, чтобы исследовать, как моторная, пространственная и визуальная информация интегрируется во время ВМО, управляемой визуальными материалами. С другой стороны, признавая ограничения традиционных методов, таких как динамическое каузальное моделирование и грайнджеровская причинность, мы применяем дата-ориентированный, нелинейный подход – многошкальную энтропию переноса – для оценки причинных сетей ЭЭГ, связанных с ВМО. Мы также рассматриваем основные эффекты руки (левая против правой) и перспективы (VMI1 против VMI3). Наконец, хотя наша основная цель заключается в выяснении активности теменно-затылочной сети во время ВМО, мы также исследуем типичные характеристики времени-частоты ВМО. Эти результаты помогут прояснить уникальную динамику мозга, лежащую в основе ВМО, и отличить ее от тех, что связаны с КМО.
Методы
Участники
В эксперименте приняли участие 17 добровольцев (3 женщины, средний возраст: 24,3 ± 1,9 года), все правши, с нормальным зрением и без анамнеза психических заболеваний. Доминирующая рука участников оценивалась с помощью Эдинбургского опросника по правшести (EHI). Средний балл составил 89,7, стандартное отклонение – 13,3. Перед экспериментом проводились тренировочные сессии, чтобы убедиться, что участники полностью понимают задачу. После эксперимента участники заполнили анкету Vividness of Movement Imagery Questionnaire2 (VMIQ2) для оценки их способности к визуально-моторной образности (ВМО). Данные от участников, получивших низкие баллы, были исключены из последующего анализа. Данное исследование проводилось в соответствии с этическими принципами Университета Кюсю и Хельсинкской декларацией. Все экспериментальные протоколы были одобрены Университетом Кюсю (ISEE 202412). Информированное согласие было получено от всех участников или их законных представителей.
Экспериментальный протокол
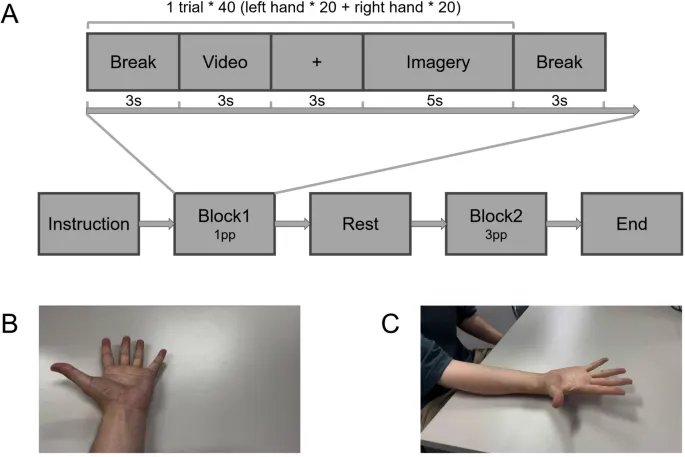
Экспериментальная процедура состояла из двух блоков. В первом блоке участники должны были выполнить задачу визуально-моторной образности с перспективой от первого лица. В частности, после паузы между пробами на экране отображалось трехсекундное видео, на котором случайным образом показывалось действие попеременного раскрытия ладони и сжатия кулака левой или правой рукой с перспективой от первого лица (1pp). Затем, после трехсекундного периода фиксации крестом, участникам предлагалось выполнить образность того же движения, что и в условии, только что показанном видео, в течение пятисекундного периода образности до появления сигнала покоя на экране. Второй блок эксперимента следовал той же процедуре, что и первый, за исключением того, что видео-инструкции и задачи образности представляли одни и те же движения рук с перспективой от третьего лица (3pp). Задачи образности случайным образом повторялись по 20 раз для каждого условия в каждом блоке, что составило в общей сложности 80 испытаний для всего эксперимента.
Во время периода образности участникам было предложено держать глаза открытыми, а руки расслабленными на бедрах. Между двумя блоками участники имели не менее 3 минут для отдыха и расслабления.
Измерение и предобработка ЭЭГ
В данном эксперименте данные ЭЭГ записывались с использованием 64-канальной системы (Nihon Kohden EEG2100) в соответствии с системой размещения электродов 10-10 с частотой дискретизации 1000 Гц. Референтный электрод был размещен на правом мочке уха. Офлайн-предобработка проводилась для данных каждого участника, включая усредненное ре-реферирование, полосовую фильтрацию (0,1–50 Гц) и удаление артефактов. В частности, использовался метод анализа независимых компонент (ICA) с 15 компонентами, частотой дискретизации 1000 Гц и алгоритмом FastICA для удаления артефактов, вызванных движениями глаз, сердечной активностью и мышечной активностью. Компоненты артефактов идентифицировались путем ручной инспекции на основе временных рядов, характеристик спектра мощности и топографических карт каждой компоненты, с использованием инструментов визуализации ICA, предоставляемых в библиотеке MNE (версия 1.6.1).
На основе распределения основных областей мозга, активированных во время ВМО, как сообщалось в предыдущих исследованиях, включая области, близкие к моторной коре, задним теменным областям и затылочным областям, были выбраны электроды C3, C4, P5, P6, O1 и O2 как электроды интереса для последующего анализа в данном эксперименте.
Событийно-связанная десинхронизация/синхронизация
ERD/ERS (событийно-связанная десинхронизация/синхронизация) часто используются для отражения тенденций в синхронизации корковой нейронной активности в интересующей области во время задачи, что определяется как процентное увеличение/уменьшение средней мощности во время события относительно базового периода:
$$RA() = eft( rac{ ext {Act} R}{R} ight) imes 100$$где Act – значение мощности в событийно-связанный период, R – средняя мощность в базовый период, а RA – относительная амплитуда. Базовый период в данном исследовании был установлен с 2 с до 1 с до начала периода образности.
Многошкальный анализ
При анализе энтропии ЭЭГ часто используется метод многошкального анализа для грубого осреднения (coarsegraining) необработанных временных рядов. Различные степени грубого осреднения позволяют нам улавливать полезную информацию на разных временных масштабах. Было показано, что многошкальный подход, основанный на усреднении точек выборки, является схемой, которая может эффективно представлять динамические различия и применим к анализу биологических временных рядов.
$$y_j^{( au )} = rac{1}{s} um _{i=(j1)s+1}^{js} x_i, uad 1 e j e rac{N}{s}$$где y_{j} – точка в грубо осредненном ряду, s – масштабный коэффициент, а x_{i} – элемент исходного временного ряда. Меньший масштабный коэффициент указывает на то, что грубо осредненный сигнал ближе к исходному сигналу, тем самым сохраняя больше деталей из исходного сигнала. В то время как больший масштабный коэффициент указывает на большее игнорирование деталей и улавливание более широких тенденций.
Реконструкция фазового пространства / символизация на основе перестановок
Реконструкция фазового пространства (PSR) является широко используемым методом для характеристики нелинейной динамики временного ряда путем реконструкции эквивалентного аттрактора в фазовом пространстве. Данный дискретный временной ряд:
$$X_k (i) = [x(i), x(i+ au ), ots , x(i+(m1) au )]$$где m – размерность вложения, обозначающая количество измерений в реконструированном фазовом пространстве. В данном исследовании m было выбрано равным 3 для баланса между сложностью системы и длиной последовательности. au – фактор задержки, который указывает интервал, через который выбираются координаты реконструированной последовательности для интерполяции. Слишком малое значение au приведет к потере информации о динамике исходной системы во время процесса реконструкции, в то время как слишком большое значение au приведет к избыточности информации во время реконструкции. В данном исследовании au определялся методом автокорреляционной функции.
После получения многомерной последовательности PSR она обычно кодируется в сочетании с методом символизации на основе перестановок для получения последовательности состояний, которая может представлять ее динамическую информацию. Для j-й компоненты реконструированной последовательности сначала упорядочиваются элементы в порядке возрастания:
$$x(i+(j_11) au ) e x(i+(j_21) au ) e ots e x(i+(j_m1) au )$$Затем извлекается порядок значений индексов каждого элемента как представление состояния текущей компоненты:
$$S_j = j_1, j_2, ots , j_m $$В данном эксперименте, поскольку размерность вложения m была выбрана равной 3, количество возможных состояний для каждой компоненты в реконструированной последовательности составляет m! = 6.
Энтропия переноса
Энтропия переноса (TE) – это непараметрический метод, основанный на теории информации, который может использоваться для эффективного расчета направленного потока информации между двумя последовательностями ЭЭГ. Следовательно, он часто используется для построения сетей связности ЭЭГ, позволяя нам получать причинные взаимосвязи различных областей мозга. TE из одного временного ряда Y в другой X определяется как:
$$T_{Y ightarrow X} = um p(x_{n+1}, x_n^{(k)}, y_n^{(l)}) og eft( rac{p(x_{n+1} id x_n^{(k)}, y_n^{(l)})}{p(x_{n+1} id x_n^{(k)})} ight)$$Очевидно, что TE из Y в X не обязательно равна TE из X в Y (обычно они неравны). Следовательно, путем расчета TE между различными интересующими каналами, мы можем получить матрицу смежности, которая представляет поток неравной информации между этими областями, что, в свою очередь, позволяет провести дальнейший анализ характеристик соответствующей сети.
Степень узла
Степень узла является ключевой характеристикой в теории графов, которая количественно определяет важность узла в сети. Обычно она определяется как количество ребер, соединенных с узлом. Поскольку сеть связности, построенная на основе энтропии переноса в данном исследовании, является направленной взвешенной сетью, степени исхода и входа определяются следующим образом:
$$d_{ ext {out}}(u) = um _{(u,v) n E} w(u,v)$$$$d_{ ext {in}}(u) = um _{(u,v) n E} w(v,u)$$где d_{out} – степень исхода узла, обозначающая сумму весов информационных потоков, исходящих из этого узла. d_{in} – степень входа узла, обозначающая сумму весов информационных потоков, входящих в этот узел. Большая степень узла обычно указывает на большее участие в сети.
Результаты
Анализ времени-частоты
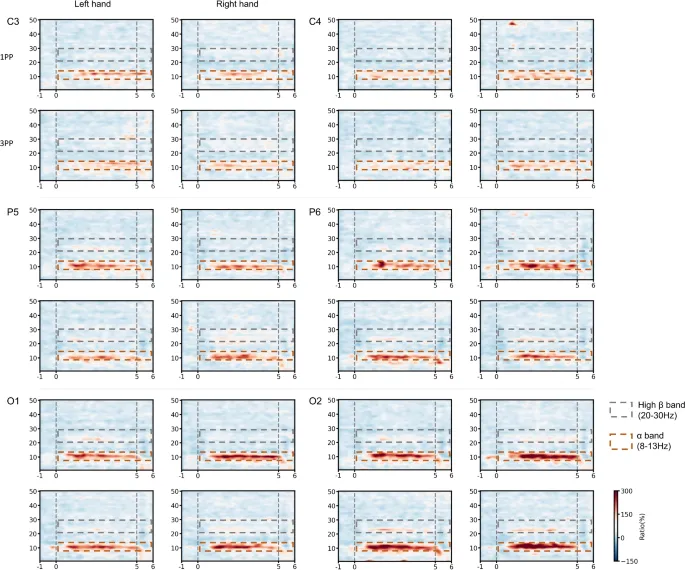
На рисунке 1 показаны ERD/ERS шести каналов. Во всех экспериментальных условиях и на всех каналах примерно через 1 секунду после начала образности наблюдалась значительная ERS в альфа-диапазоне (8–13 Гц), которая сохранялась до конца выполнения задачи. Дополнительно наблюдалась относительно слабая ERS в высокобета-диапазоне (20–30 Гц).
Стоит отметить, что ни одно из вышеупомянутых сравнений между группами усредненных ERS не показало статистически значимых различий (тест Уилкоксона на знаковые ранги, p<0,05).
Сеть связности
Многошкальная символизация на основе перестановок
Многошкальный анализ проводился для двух частотных диапазонов, выявленных в анализе времени-частоты. Выбор масштабного коэффициента основывался на двух соображениях. Во-первых, он должен был гарантировать, что длина грубо осредненных последовательностей соответствовала требованиям для последующих расчетов PSR и TE. Во-вторых, он должен был быть соответствующим с точки зрения физического смысла PSR. В данном исследовании фактор задержки PSR определялся методом автокорреляционной функции. При увеличении масштабного коэффициента фактор задержки уменьшался до 1. Если масштабный коэффициент был слишком велик, даже при факторе задержки 1 (что приводило к очень компактной реконструкции), существовал риск потери существенной динамической информации исходной системы.
Для отражения различий TE на разных масштабах мы выбрали масштабные коэффициенты 4 и 16 для альфа-диапазона, охватывающие как малые масштабы, сохраняющие больше деталей, так и большие масштабы, фильтрующие избыточные детали. Для высокобета-диапазона мы выбрали масштабные коэффициенты 3 и 8, следуя тому же принципу. Далее мы применили символизацию на основе перестановок к грубо осредненным последовательностям в обоих частотных диапазонах, получив последовательности состояний, необходимые для расчетов TE.
Энтропия переноса
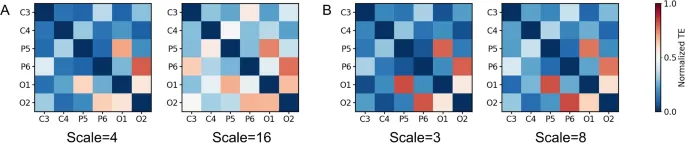
Для шести интересующих каналов были вычислены матрицы смежности при различных масштабных коэффициентах. В этих матрицах элементы представляют собой связи между различными каналами, а их цвета соответствуют нормализованным значениям TE, усредненным по всем участникам.
Сети связности предполагали общие закономерности по всем четырем масштабам и обоим частотным диапазонам. Во-первых, они последовательно показывали доминирующий поток информации между двумя парами однополушарных областей мозга: P5–O1 и P6–O2. Во-вторых, значительный поток информации также наблюдался между каналами O в разных полушариях, а также между каналами P/O и их контралатеральными каналами C. На масштабе 4 в альфа-диапазоне и масштабах 3 и 8 в высокобета-диапазоне поток информации в P5–O1 и P6–O2 был особенно доминирующим. Однако на большем масштабе (масштаб 16, альфа) доля другого информационного потока увеличилась.
Анализ степеней узлов сети
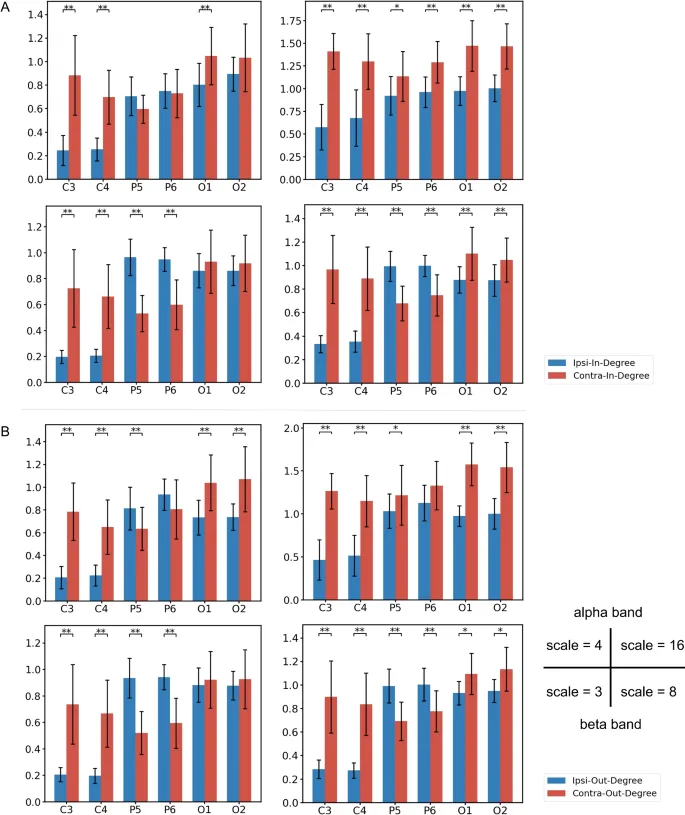
Анализ степеней узлов сети далее раскрыл закономерности информационных потоков, сосредоточенных вокруг каждого канала. Для C3 и C4 наблюдались значительные контралатерально-доминирующие входящие и исходящие потоки во всех условиях. Для P5 и P6 контралатеральный поток информации превосходил ипсилатеральный поток в альфа-диапазоне на масштабе 16, в то время как другие условия показывали ипсилатерально-доминирующую модель переноса. Для O1 и O2 контралатеральный информационный перенос преобладал во всех условиях.
Эффект руки
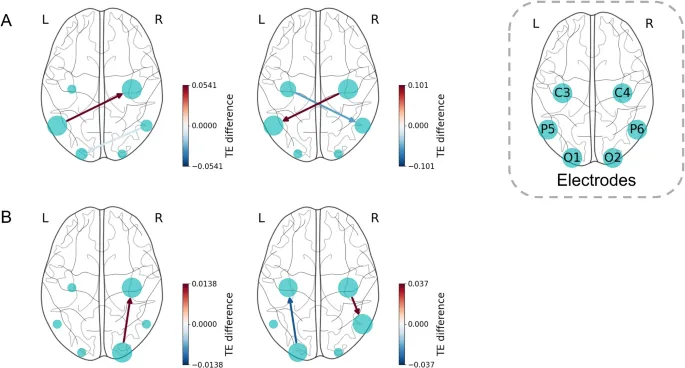
Сравнение сетей TE между экспериментальными условиями выявило значительные различия в информационном потоке в зависимости от эффектов руки и перспективы (N=34, тест Уилкоксона на знаковые ранги).
В альфа-диапазоне на обоих масштабах существуют информационные потоки, где группа левой руки значительно превосходила группу правой руки между P5 и C4. Примечательно, что на малом масштабе доминирующее направление информационного потока было P5 → C4, тогда как на большом масштабе – C4 → P5. Кроме того, на масштабе 4, P6 → O1 показывал более слабый информационный поток в группе левой руки, чем в группе правой руки. На масштабе 16 наблюдалась аналогичная тенденция для C3 → P6.
В высокобета-диапазоне информационный поток между C4 и его ипсилатеральной теменно-затылочной областью был больше в группе левой руки. Дополнительно наблюдались значительные информационные потоки от O2 → C4 на масштабе 3 и от C4 → P6 на масштабе 8. Более того, информационный поток O1 → C3 был значимым на масштабе 8.
Эффект перспективы
В альфа-диапазоне на масштабе 4 значимых различий в информационном потоке между группами не наблюдалось. В отличие от этого, на масштабе 16 передне-задний информационный поток был значительно больше в группе 1pp по сравнению с группой 3pp. Это включало информационные потоки от билатеральных узлов C и P к ипсилатеральным узлам O, C4 → O1, и двунаправленные потоки между C3 и P6.
В высокобета-диапазоне группа 3pp демонстрировала значительно больший информационный поток, чем группа 1pp, в правой теменно-затылочной области. На масштабе 3 наблюдались двунаправленные потоки между P6 → O1 и P6 → O2, а также от C3 → P5. На масштабе 8 значимые информационные потоки были P6 → O1 и P6 → O2.
Обсуждение
Исследования активности мозга во время ВМО были относительно ограничены, поскольку парадигмы КМО использовались в качестве шаблонов. Однако многочисленные исследования указывали на критическую роль задних теменной и затылочной областей в кодировании моторных намерений и участии в визуальной образности, что предполагает потенциал применения ВМО в дальнейших разработках. В данном исследовании мы сначала подтвердили характеристики спектра мощности ВМО с использованием анализа времени-частоты, широко используемого метода для захвата макроскопических паттернов ЭЭГ МО. Основываясь на этом, мы исследовали направленную связность в соответствующих частотных диапазонах. Впервые был использован подход на основе многошкальной энтропии переноса для исследования эффективной сети связности ЭЭГ моторной образности.
В отличие от хорошо известных характеристик времени-частоты ЭЭГ КМО, консенсус в отношении ЭЭГ ВМО до сих пор отсутствует. Эта предвзятость может быть связана с вариациями в экспериментальных парадигмах и спонтанной активностью ЭЭГ у участников, что затрудняет контроль всех влияющих факторов. Более того, обычная модель, в которой КМО подавляет нейронную синхронизацию в моторной коре, может быть неприменима напрямую к ВМО. Хотя некоторые исследования сообщали о явлениях ERD в теменной или затылочной долях во время ВМО, наше исследование не обнаружило таких эффектов. Это расхождение может быть объяснено различиями в сенсорных модальностях, подчеркнутых экспериментальным дизайном. В частности, степень, в которой участники воспринимают образность как «моторные ощущения» или «визуальные переживания», может влиять на нейронную активность. Первое, вероятно, задействует существующие моторные шаблоны (связанные с двигательным исполнением), в то время как второе больше соответствует визуальному восприятию (связанному с наблюдением за движением) и содержит меньше моторно-перцептивных компонентов. Учитывая, что наше исследование использовало видео в качестве направленного стимула, сигналы ЭЭГ из теменно-затылочных областей во время образности демонстрировали последовательную ERS. Это предполагает, что подчеркивание явных визуальных сенсорных атрибутов в задаче эффективно изолирует кинестетические ощущения от моторной образности.
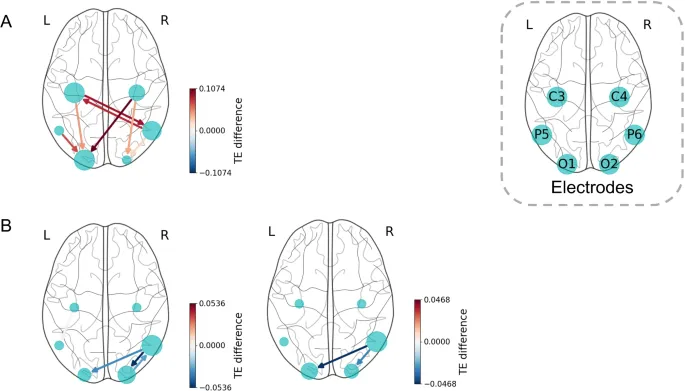
Наш сетевой анализ подтверждает предыдущие выводы о том, что задние теменные и затылочные области, а не моторная кора, играют доминирующую роль в ВМО. На макроскопическом уровне процесс ВМО в основном характеризовался сильным потоком информации между ипсилатеральными нижними теменными и затылочными долями, а также билатеральными затылочными областями. Дополнительно наблюдались двунаправленные связи между теменными и моторными областями (преимущественно контралатеральные) на различных масштабах.
Интеграция многошкального анализа с PSR выявила новые представления о направленной связности, включая ранее не сообщавшиеся явления. В анализе эффекта руки мы обнаружили последовательные межгрупповые различия как в альфа-, так и в высокобета-диапазонах. Однако направленные связи различались в зависимости от масштаба: C4 → P5 в альфа-диапазоне и C4 → P6/O2 в высокобета-диапазоне. Это предполагает различную временную зависимость в обмене информацией между этими областями, которую трудно уловить с помощью обычных методов символизации, применяемых к однородным временным рядам.
Кроме того, направленная связность, включающая каналы моторной области C3 и C4, следовала четкому контралатеральному паттерну и демонстрировала значительный эффект руки. Однако аналогичный паттерн не наблюдался для эффекта перспективы. Примечательно, что в альфа-диапазоне этот эффект был ограничен взаимодействиями между моторными и задними теменными областями и не распространялся на затылочные области. В высокобета-диапазоне он включал прямую связность между моторными и затылочными областями. Эти результаты указывают на то, что эффект руки модулирует визуальное/пространственное внимание в задних теменных и затылочных областях через процессы, связанные с моторикой, но не влияет на базовое возбуждение зрительной коры.
Что касается анализа эффекта перспективы, то в альфа-диапазоне образность с перспективой 1pp демонстрировала значительно больший обратный поток информации, особенно в направлении затылочной области, по сравнению с образностью 3pp. Хотя результаты VMIQ2 не указывали на различия в субъективной яркости двух перспектив (1pp: 1,86 ± 0,74, 3pp: 1,81 ± 0,63), анализ направленной связности выявил усиленную нисходящую модуляцию, сосредоточенную в затылочной доле для образности 1pp. Этот процесс включает прямое и косвенное (через заднюю теменную долю) модуляцию, связанную с задачей, из моторной области, наряду с пространственной сенсорной регуляцией из задней теменной области, совместно усиливающую генерацию визуального опыта в затылочной коре.
В высокобета-диапазоне образность 3pp показывала более сильный информационный поток из правой задней теменной доли в затылочную долю по сравнению с образностью 1pp. Это предполагает уникальную роль правой задней теменной области в модуляции активности затылочной коры с внешней точки зрения через пространственную локализацию, основанную на внимании.
Данное исследование предоставляет новые данные об эффективной связности ЭЭГ ВМО, демонстрируя доминирующую роль задних теменной и затылочной областей в обработке моторной образности. Применяя многошкальный подход энтропии переноса, мы выявили уникальные паттерны связности, зависящие как от эффекта руки, так и от эффекта перспективы, предполагая лежащие в основе механизмы для различных типов стратегий моторной образности. Эти результаты способствуют более глубокому пониманию нейронной динамики ВМО, но следует уточнить несколько ограничений.
Во-первых, хотя основной целью данного исследования было изучение направленных взаимодействий между конкретными областями интереса (ROI), мы также должны подчеркнуть важность открытия крупномасштабной сетевой динамики, как все чаще сообщается в недавних исследованиях связности всего мозга. Наш фокус был направлен на захват мелкомасштабных модуляций в областях, наиболее тесно связанных с ВМО, и построение, а также анализ полной сети 64 × 64 направленных связей было бы не только вычислительно затратным, но и менее практичным для прикладных интерфейсов ЭЭГ. Следовательно, мы применили гипотезо-ориентированный подход к области интереса (ROI), нацеленный на зрительные, теменные и моторные области, ранее вовлеченные в литературу по ВМО. Будущие исследования, сочетающие ЭЭГ высокой плотности с одновременной фМРТ, могут расширить эти выводы до истинно крупномасштабных паттернов координации областей всего мозга.
Во-вторых, из-за потенциальной предвзятости, вносимой невысокой плотностью ЭЭГ в анализе на уровне источников, данное исследование использовало подход на уровне масштабов для вычислений и анализа. Хотя этот метод сохраняет некоторый риск утечки источников из-за объемного проведения ЭЭГ, сочетание символизации на основе перестановок и ненулевых задержек вложения (τ = 816 мс) эффективно подавляет связи, возникающие из-за связи с нулевой задержкой. Кроме того, использование общего среднего референса помогает смягчить предвзятость, связанную с референсом.
Наконец, как обсуждалось ранее, данное исследование использовало экспериментальный протокол, который явно ограничивал шаблоны образности участников. Хотя этот подход помог контролировать экспериментальные переменные, он также упустил из виду вариативность в индивидуальных стратегиях образности. В реальных приложениях моторной образности такие факторы, как ясность целевого направления, сенсорные атрибуты, подчеркиваемые во время образности, и предыдущий опыт участника с действием, могут влиять на стратегии образности, приводя к различным паттернам мозговой активности. Следовательно, дальнейшее исследование независимой от задачи и обобщенной мозговой связности во время ВМО необходимо для повышения применимости ЭЭГ ВМО в практических сценариях.
Данные
Представленные в данном исследовании данные доступны по запросу у соответствующего автора. Данные не находятся в открытом доступе из-за ограничений конфиденциальности участников.
Ссылки
[Ссылки на литературу отсутствуют в предоставленном тексте, но в реальном исследовании они были бы здесь.]
Информация об авторах
Аспирантура и факультет информационных наук и электротехники, Университет Кюсю, Фукуока, 8190382, Япония
- Хаобо Чжоу
- Кейджи Ирамина
Авторство
- Хаобо Чжоу: Просмотр публикаций автора на PubMed, Google Scholar
- Кейджи Ирамина: Просмотр публикаций автора на PubMed, Google Scholar
Вклад
Концептуализация и предварительный эксперимент: Х.З., К.И., Реализация эксперимента: Х.З., Анализ данных и визуализация: Х.З., Написание и редактирование: Х.З. и К.И., Руководство: К.И. Все авторы ознакомились с рукописью.
Соответствующий автор
Корреспонденция с Хаобо Чжоу.
Заявления об этике
Конфликты интересов
Авторы заявляют об отсутствии конкурирующих интересов.
Дополнительная информация
Примечание издателя
Springer Nature остается нейтральной в отношении юрисдикционных претензий в опубликованных картах и институциональной принадлежности.
Права и разрешения
Открытый доступ
Данная статья лицензирована в соответствии с Международной лицензией Creative Commons Attribution 4.0, которая разрешает использование, распространение, адаптацию, переработку и воспроизведение на любом носителе или в любом формате, при условии, что вы должным образом указываете авторство и источник, предоставляете ссылку на лицензию Creative Commons и указываете, были ли внесены изменения. Изображения или другие материалы третьих сторон, включенные в статью, относятся к лицензии Creative Commons статьи, если не указано иное в подписи к материалу. Если материал не включен в лицензию Creative Commons статьи, и предполагаемое вами использование не разрешено законодательством или превышает разрешенное использование, вам необходимо получить разрешение непосредственно у правообладателя. Чтобы просмотреть копию данной лицензии, посетите http://creativecommons.org/licenses/by/4.0/.
Перепечатки и разрешения
Об этой статье
Цитировать эту статью: Zhou, H., Iramina, K. Discovery of EEG effective connectivity during visual motor imagery with multiscale symbolic transfer entropy. Sci Rep 15, 38200 (2025). https://doi.org/10.1038/s41598025221437
Скачать цитату
Получено: 17 марта 2025 г.
Принято: 25 сентября 2025 г.
Опубликовано: 31 октября 2025 г.
DOI: https://doi.org/10.1038/s41598025221437
Поделиться этой статьей
Любой, кто поделится по следующей ссылке, сможет прочитать этот контент:
Получить ссылку для обмена
К сожалению, ссылка для обмена в настоящее время недоступна для этой статьи.
Копировать ссылку для обмена в буфер обмена
Предоставлено инициативой Springer Nature SharedIt по обмену контентом.
Ключевые слова
Моторная образность, ЭЭГ, Сеть связности, Энтропия переноса





