Интересное сегодня
Природа цветовых послеобразов: научное исследование адаптации зрения
Введение: Как возникают цветовые послеобразы
Цветовой послеобраз возникает, когда наблюдатель в течение длительного времени фокусирует взгляд на цветной области. В результате этого воздействия, лежащие в основе механизмы обработки цвета адаптируются и становятся нечувствительными к цвету этой области, индуктору. Эти механизмы остаются наиболее чувствительными к цвету, который наименее похож на индуктор, и производят комплементарный (или негативный) послеобраз при замене индуктора бесцветной серой областью. Например, если вы сосредоточитесь на желтом круге, после его исчезновения вы увидите сине-пурпурный круг, даже если площадь круга теперь физически совпадает с бесцветным окружением (см. Дополнительное видео 1 и раздел A.1 Иллюстрация послеобразов Дополнительных материалов).
Основные механизмы временной адаптации к цвету позволяют нам надежно воспринимать цвет, несмотря на постоянные изменения освещения в нашей повседневной среде. Иллюзорные послеобразы, таким образом, использовались для распространения самых фундаментальных принципов человеческого цветового зрения в обзорах, учебниках и других более широких коммуникациях, включая концепции хроматической адаптации и цветовой постоянства, цветовой оппонентности и комплементарности, а также зависимости цветов от контекста и наблюдателя.
Противоречия и запутанность в объяснениях послеобразов
Несмотря на то, что возможные механизмы адаптации известны более века, современные объяснения этого важного явления остаются противоречивыми и запутанными. Человеческое цветовое зрение является результатом нескольких стадий обработки, начиная с возбуждения колбочек в сетчатке, распространяясь через колбочно-оппонентные механизмы в ганглиозных клетках сетчатки, латеральном коленчатом теле таламуса и одноклеточных и двухклеточных оппонентных клетках первичной зрительной коры, пока цветовой сигнал не достигнет высших кортикальных областей, которые производят субъективное ощущение цвета.
Свидетельства адаптации были обнаружены на многих этапах этой визуальной иерархии. Адаптация фоторецепторов на первом этапе цветовой обработки производит дивергентную адаптацию, подобную первоначальному предложению фон Криса, и может быть приближена кодированием контраста по закону Вебера. Более медленный тип «вторичной адаптации» происходит в ганглиозных клетках сетчатки и предполагается субтрактивным. Другие наблюдения предполагали «высшую» адаптацию кортикальных механизмов.
Научные дебаты о природе послеобразов
Исследователи расходятся во мнениях относительно нейронных механизмов адаптации, вызывающих комплементарные послеобразы. Классические исследования предполагали, что такие послеобразы вызваны десенсибилизацией фоторецепторов («выцветание колбочек») на первом этапе обработки. Более поздние исследования решительно опровергли эту идею, приписав послеобразы адаптации в колбочно-оппонентных каналах второго этапа, или подчеркнули эффекты кортикальной адаптации.
Еще более важно, что связь между нейронными механизмами и воспринимаемыми цветами послеобразов игнорировалась или искажалась. Независимо от предполагаемого нейронного происхождения, большинство современных исследований предполагает, что воспринимаемые цвета послеобразов являются колбочно-оппонентными, но существуют и доказательства, опровергающие это предположение. В результате этой путаницы учебники, обзоры и другие более широкие коммуникации либо не пытаются объяснить цветовые послеобразы, либо распространяют вводящие в заблуждение нарративы, такие как идея, что послеобразы отражают херенговскую оппонентность между красным/зеленым и синим/желтым.
Данное исследование: новый взгляд на цветовые послеобразы
Данное исследование использовало тот факт, что адаптация механизмов на разных этапах обработки цвета дает значительно разные количественные предсказания относительно цветов комплементарных послеобразов (Рис. 1). Предсказываемые оттенок и насыщенность послеобразов различаются в разных моделях. Оттенок описывает, насколько красноватым, желтоватым, зеленоватым и синеватым является цвет. Насыщенность (Chroma) относится к яркости цвета оттенка. При равной яркости (изолюминансности) она эквивалентна насыщенности и соответствует отличию цвета от серого.
С помощью специально разработанных экспериментальных парадигм были измерены точные цвета, воспринимаемые в послеобразах, для широкого диапазона индукторов, чтобы проверить эти предсказания. Три всесторонних эксперимента недвусмысленно показали, что цвета послеобразов тесно следуют неоппонентным предсказаниям, специфичным для адаптации колбочек.
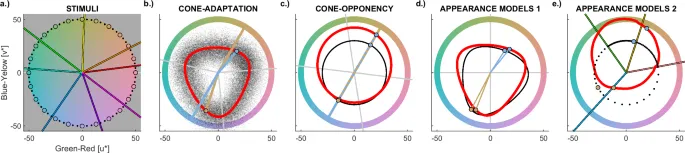
Рис. 1: Стимулы послеобразов и модели
Во всех панелях цвета представлены в цветовом пространстве CIELUV; см. Рис. S11 Дополнительных материалов для аналогичных диаграмм в колбочно-оппонентном пространстве DKL. a иллюстрирует образцы стимулов в Эксперименте 1a (цветные линии), Эксперименте 1b (черные круги) и Эксперименте 2a (черные круги и точки). В Эксперименте 2b стимулы были выбраны вдоль цветового круга, как в Эксперименте 2a, но в DKL, а не в CIELUV (Рис. S11.a). b показывает результаты адаптации колбочек без (красная линия) и с шумом (черные точки); шум использовался для моделирования гистограмм оттенков. c иллюстрирует адаптацию колбочно-оппонентных каналов, где послеобраз смещен к оттенку, противоположному оттенку индуктора (толстая красная линия). Для сравнения также показаны предсказания, основанные на адаптации в пространстве CIELUV (тонкая черная линия). Предсказания оттенков CIELUV такие же, как и для колбочно-оппонентного пространства DKL, но предсказываемая насыщенность отличается от колбочно-оппонентных предсказаний. d иллюстрирует предсказания CIECAM02 (толстая красная линия) и CIELAB (тонкая черная линия), которые являются распространенными моделями цветового зрения. e иллюстрирует херенговскую оппонентность (черный) и оппонентность в системе цветов Манселла (красный). Цветные линии на фоне соответствуют измеренным красным, желтым, зеленым и синим. Два колбочно-оппонентных примера индукторов (60 и 240°) показаны на (b–e). Точка пересечения линий и цветового круга указывает на индуктор, большие точки указывают на цвет соответствующего послеобраза, предсказанный соответствующей моделью. Линии и точки показаны цветом индуктора. Для личного восприятия желтоватый индуктор (60°) соответствует индуктору в Дополнительном фильме 1.
Методы: Экспериментальный дизайн
Эксперимент 1 был одобрен этическим комитетом Университета Гиссен (LEK 20170030). Эксперименты 2 и 3 были одобрены Этическим комитетом факультета Университета Саутгемптона, ERGO 65442. Все участники предоставили информированное согласие перед участием. Ни один из экспериментов не был предварительно зарегистрирован. Вся информация о поле предоставляется участниками.
Эксперимент 1: Послеобразы фиксированного положения
Участники
В Эксперименте 1a приняли участие 31 наблюдатель (25 женщин и 6 мужчин, возраст: M = 25.9 ± SD = 4.2 года). В Эксперименте 1b приняли участие 52 наблюдателя (36 женщин и 16 мужчин, возраст: 25.1 ± 4.3 года). Участники были вознаграждены зачетными единицами или 8 евро в час. Дефекты цветового зрения были исключены с помощью таблиц HRR.
Аппаратура
Стимулы представлялись на мониторе Eizo Colour Edge (36.5 × 27 см) с графической картой AMD Radeon Firepro с разрешением цвета 10 бит на канал. Координаты цветности CIE1931 и яркость (xyY) первичных цветов монитора были: R = (0.6847, 0.3111, 26.4), G = (0.2138, 0.7263, 69.9), B = (0.1521, 0.0453, 4.8). Гамма составляла 2.2 для всех каналов и была скорректирована.
Стимулы
Цвета были представлены в пространстве CIELUV. Точка белого была xyY = [0.3304, 0.3526, 101.1], яркость фона L* = 70. При изолюминансности оппонентные оттенки в CIELUV совпадают с колбочно-оппонентными оттенками в пространстве Derrington-Krauskopf-Lennie (DKL) (Рис. 1a) и с оппонентными оттенками, определенными вдоль линий в (гамма-корректированном) пространстве HSV и RGB, значениями тристимулуса (XYZ) и координатами цветности (xyY); см. раздел E.23 Дополнительных материалов для математических деталей. Пространство CIELUV было предпочтительнее DKL, поскольку оно лучше контролирует воспринимаемую насыщенность.
Восемь цветов индукторов в Эксперименте 1a были выбраны так, чтобы соответствовать типичной яркости и оттенку красного, оранжевого, желтого, зеленого, бирюзового, синего, фиолетового и пурпурного при максимальной возможной насыщенности в пределах гаммы монитора (цветные линии на Рис. 1a). Девять оттенков сравнительных цветов (см. Процедура) были получены путем добавления четырех оттенков с шагом 10 градусов (градусов) в любом направлении (низкий или высокий азимут) от оппонентного оттенка. Яркость и насыщенность сравнений были определены в результате предварительных испытаний.
В Эксперименте 1b двадцать четыре индуктора были выбраны вдоль цветового круга в CIELUV с насыщенностью 71 и равными шагами 15 градусов, начиная с 0 градусов (черные круги на Рис. 1a). Насыщенность была выбрана как максимальная, достижимая в пределах гаммы монитора для всех направлений оттенков. Яркость индукторов, сравнительных цветов и ахроматического диска в центре тестового дисплея была такой же, как и у фона (L* = 70). Эта яркость соответствует 1.7% выцветания колбочек согласно константе полувыцветания Раштона и Генри. Оттенки сравнительных цветов определялись так же, как и в Эксперименте 1a, а насыщенность была сохранена постоянной на уровне 30 для всех сравнительных цветов. Этот уровень насыщенности был определен в результате предварительных испытаний и учитывает более низкую насыщенность послеобразов по сравнению с индукторами. Таблица S2 Дополнительных материалов содержит подробные спецификации цветов.
Процедура
В каждом испытании индукторный дисплей (Рис. 2a) показывался в течение 20 секунд (Эксперимент 1a) и 30 секунд (Эксперимент 1b). Задачей-прикрытием гарантировалось, что наблюдатель фиксировал точку в центре (подробности в разделе B.2 Подробности измерений послеобразов Дополнительных материалов). После этого периода адаптации наблюдатели определяли, какой из девяти сравнений был наиболее близок к цвету послеобраза, видя послеобраз в центре (Рис. 2a). Послеобразы, казалось, «сливались» с сегментом, цвет которого был наиболее близок к послеобразу, делая задачу интуитивно понятной. Участники использовали мышь, чтобы указать сегмент с совпадающим сравнением. Если они не видели ни одного из сравнительных цветов в центре, они могли также щелкнуть по центральному диску, чтобы указать на это и пропустить испытание (результат - пропущенное значение).
Использовался 10-секундный межпробный интервал, за которым следовал самостоятельно назначаемый перерыв, чтобы отменить оставшиеся послеобразы и предотвратить их интерференцию между испытаниями (см. раздел B.2 Подробности измерений послеобразов Дополнительных материалов). Порядок испытаний был рандомизирован. В Эксперименте 1a испытания повторялись в четырех блоках. Несколько испытаний были пропущены (около 2% в обеих частях).
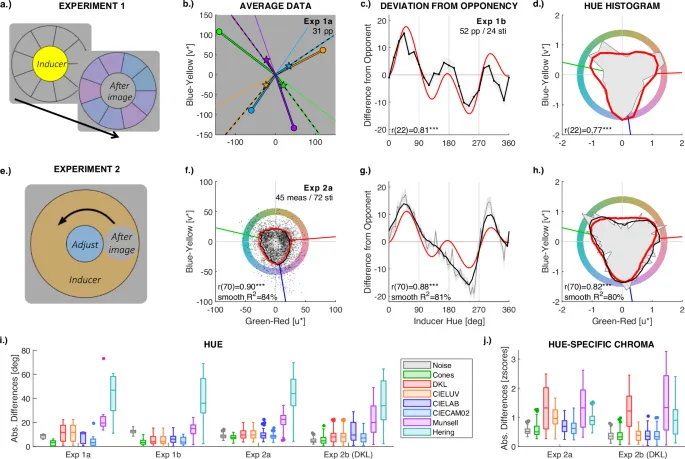
Рис. 2: Эксперимент 1 и 2
Первый ряд (a–d) иллюстрирует стимульные дисплеи и задачу (a), а также результаты (c, d) для Эксперимента 1. Наблюдатели адаптируются к индукторному дисплею в течение 30 секунд (желтый на a). Затем индукторный дисплей заменяется кругом фонового серого цвета вместе с девятью сравнительными цветами (пурпурный на a) в круговой схеме вокруг серого круга. Серый круг в центре воспринимается как индуцированный послеобраз, и наблюдатели выбирают сравнительный цвет, который лучше всего соответствует цвету, который они видят в центре. b приводит примеры совпадений послеобразов, усредненных по N = 31 участнику в Эксперименте 1a с максимально насыщенными цветами. Диски соответствуют индукторам, звезды – средним совпадениям воспринимаемых послеобразов. Тонкая линия на фоне обозначает колбочно-оппонентное направление, пунктирная линия – предсказание модели адаптации колбочек (см. рис. S2a и S3 для результатов с другими четырьмя цветами). c, d иллюстрируют результаты от N = 52 участников в Эксперименте 1b. Второй ряд (e–h) соответствует Эксперименту 2a с парадигмой «гоняющегося» изображения. На (e) хроматическое кольцо является индуктором. Участники фиксируют центр, и движущийся серый круг на кольце выявляет послеобраз. Наблюдатели настраивают оттенок и насыщенность центрального круга, чтобы он соответствовал движущемуся. Средние совпадения послеобразов от N = 10 участников и общие N = 45 измерений на цвет (черная кривая на f) тесно соответствуют предсказаниям адаптации колбочек (красный контур), давая корреляцию (внизу слева) между измеренной и смоделированной интенсивностью послеобраза (т.е. насыщенностью) по всем N = 72 индукторным цветам. Кривые в третьей колонке (c, g) показывают отклонение модели адаптации колбочек (красная кривая) и измеренных послеобразов (черная кривая) от цветов, оппонентных индукторам. Корреляции между смоделированными и измеренными отклонениями от оппонентности показаны внизу диаграмм. Корреляция отражает высокое сходство профилей двух кривых. Гистограммы оттенков в последней колонке (d, h) подсчитывают отклики оттенков (азимут на f) и отображают полученные частоты в зависимости от азимута в полярном графике. Гистограмма измерений показана серым фоном, а гистограмма смоделированных послеобразов – красным контуром. Панель d предоставляет гистограмму оттенков для N = 52 измерений для 24 цветов в Эксперименте 1b, а (h) – для N = 45 измерений для 72 индукторных цветов в Эксперименте 2a. На (h) сглаженная версия гистограммы оттенков показана черной линией. Гистограммы оттенков имеют три кластера, которые тесно соответствуют предсказаниям модели (красная линия), тем самым подтверждая результаты (c, f, g). Дополнительные результаты Эксперимента 2b со стимулами в пространстве DKL приведены на Рис. S12 Дополнительных материалов. Последний ряд иллюстрирует средние отклонения различных моделей предсказаний от измеренного оттенка послеобраза (i) и насыщенности (j). Верхний и нижний края каждого ящика – это верхняя и нижняя квартили, линия внутри каждого ящика – медиана; усы – минимальные и максимальные значения, которые не являются выбросами, а точки за пределами усов – это выбросы, идентифицированные как значения вне межквартильного диапазона (ящика) в 1.5 раза больше размера межквартильного диапазона. Серые символы (Шум) дают оценки шума, рассчитанные как среднее различие измерений каждого индивида от средней группы (межиндивидуальная вариация) в Эксперименте 1 и как среднее различие каждого отдельного измерения от среднего значения каждого участника (внутрииндивидуальная вариация) в Эксперименте 2. Насыщенность была стандартизирована (z-scored) для фокусировки на относительных различиях между оттенками. Дополнительные фильмы 1–4 визуализируют различия между оппонентными и послеобразными оттенками (см. также раздел A и Рис. S1 Дополнительных материалов).
Цветовое именование и контрольная задача
Подробности измерения категорий и прототипов цветов, а также контрольной задачи см. в разделах B.3–4 Дополнительных материалов.
Эксперименты 2-3: Послеобразы типа «гоняющегося» изображения
Таблицы S3 и S4 Дополнительных материалов содержат подробную информацию об участниках и аппаратуре в Экспериментах 2 и 3. Дополнительные фильмы 1–4 и раздел A.2 Иллюстрация метода в Экспериментах 2–3 Дополнительных материалов содержат упрощенную иллюстрацию задач в Экспериментах 2 и 3.
Участники
Десять добровольцев-участников (6 женщин и 4 мужчины, возраст: M = 23.4 ± SD = 8.3 года), включая автора (CW, мужчина, 43 года), приняли участие (см. Таблицу S4) в Эксперименте 2a. Еще три наблюдателя начали один раунд измерений (8 оттенков), но не вернулись, чтобы завершить достаточный набор данных для всех 72 оттенков. CW и одна наивная наблюдательница 37 лет (f1) участвовали в Эксперименте 2b (см. Таблицу S4). В Эксперименте 3 измерения по насыщенности проводились автором (CW) и четырьмя наивными 19-летними участницами (Таблица S5).
Аппаратура и стимулы
В Экспериментах 2–3 использовались три различные экспериментальные установки, откалиброванные так же, как и в Эксперименте 1 (Таблицы S4–S5). Все измерения проводились в полной темноте. На рисунке 2e показан стимульный дисплей (см. также Дополнительные фильмы 1–4 и Рис. S1). В Эксперименте 2a 72 индукторных цвета были выбраны с шагом 5 градусов по оттенку вдоль изолюминансного цветового круга при L* = 70 в пространстве CIELUV. Это разрешение выбора почти исчерпывающее, учитывая, что пороги различения оттенков больше 5 градусов. Яркость соответствовала примерно 3–3.5% выцветания колбочек согласно константе полувыцветания Раштона и Генри. Насыщенность индуктора была установлена на максимальную в пределах соответствующей гаммы монитора, что привело к насыщенности 38 (один участник), 42 (два участника) или 50 (все остальные); подробности см. в Таблице S4. В Эксперименте 2b выбор был сделан вдоль изолюминансного цветового круга в пространстве DKL с радиусом 0.5 (CW) и 0.7 (f1), как показано на Рис. S11 Дополнительных материалов. Хроматические оси были масштабированы относительно гаммы монитора, чтобы избежать артефактов гаммы (см. раздел E.2 Модель колбочно-оппонентности (DKL) Дополнительных материалов для математических деталей).
Для измерений по насыщенности в Эксперименте 3 (Рис. 3) были измерены дополнительные уровни насыщенности при 20, 30 и максимальном в пределах гаммы (см. Таблицу S5 для подробностей).
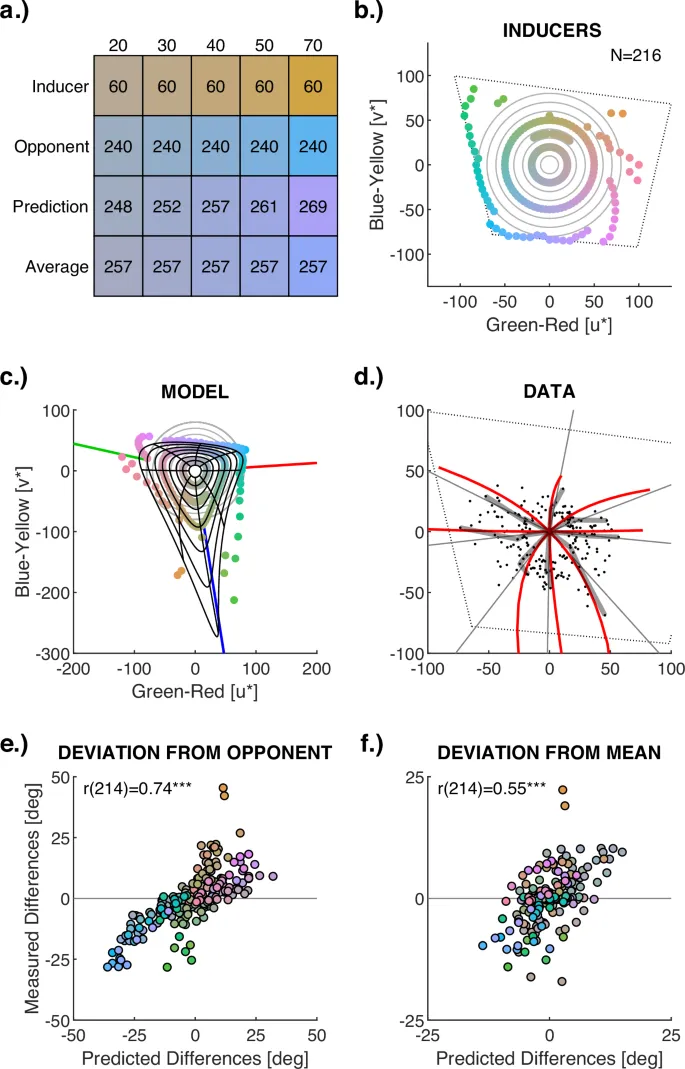
Рис. 3: Изменение оттенка послеобраза в зависимости от насыщенности (Эксперимент 3)
a Визуализация эффекта насыщенности индуктора на оттенок послеобраза. Числа в цветных прямоугольниках указывают оттенок. Строки соответствуют примерам индукторных стимулов с постоянным оттенком (60 градусов) и различными уровнями насыщенности (20–70), колбочно-оппонентным оттенком (240 градусов), предсказанием адаптацией колбочек с изменяющимся оттенком и средним оттенком предсказания, используемым для анализа. Изменение насыщенности, предсказанное адаптацией колбочек, здесь игнорируется для облегчения визуального сравнения изменений оттенка. b 216 индукторных стимулов (цветные точки), варьирующихся по трем уровням насыщенности (20, 50, максимальная). Серые круги на фоне обозначают насыщенность (радиус), варьирующуюся от 10 до 80 с шагом 10. Пунктирный тетрагон показывает гамму монитора на этом уровне яркости. c Послеобразы, смоделированные с помощью контрастов колбочек. Черные линии иллюстрируют, как послеобразы меняются, когда насыщенность индуктора соответствует серым кругам. Цветные точки обозначают смоделированные послеобразы для измерений с 216 индукторными стимулами (b). d Сравнение измерений и предсказаний адаптации колбочек. Серые точки – 216 измерений; толстые прозрачные черные линии выделяют изменение по насыщенности для оттенков с интервалом 45 градусов. Серые линии показывают эталонное среднее, используемое для расчета цветоспецифических различий по насыщенности. Красные линии показывают соответствующие предсказания адаптации колбочек. e, f Диаграммы рассеяния, иллюстрирующие корреляцию между смоделированными и измеренными изменениями оттенка послеобраза по насыщенности. Оси указывают разницу оттенка послеобраза от колбочно-оппонентного оттенка (e) и от эталонного среднего (f) в градусах азимута. Цвет точек указывает индукторные цвета. Эта фигура показывает результаты для наблюдателя f7. Результаты для четырех других наблюдателей можно найти в Таблице 4 и на Рис. S13, каждый из которых воспроизводит эти результаты.
Процедура
В каждом испытании участникам предлагалось фиксировать центр дисплея до тех пор, пока движущийся круг не достигнет максимальной цветовой насыщенности. Затем они использовали клавиши со стрелками для регулировки оттенка (влево/вправо) и насыщенности (вверх/вниз) центрального круга. Начальный цвет этого круга был установлен на случайный оттенок при насыщенности индуктора. Во время грубой настройки участники могли непрерывно изменять цвета; но перед подтверждением настроенного цвета участники должны были выполнить точную настройку (удерживая пробел). При точной настройке цвет изменяется отдельными шагами (по радиусу или по азимуту 1 градус) при каждом нажатии клавиши.
Поскольку настройка занимает время, наблюдатели видели послеобразы от настроенного цвета, которые уменьшали воспринимаемую насыщенность настроенного цвета. Были приняты две меры для избежания этого: Во время грубой настройки черные и белые круги (не области) расширялись от центра настроенного диска до его обода. Эти круги исчезали во время точной настройки, чтобы избежать интерференции с окончательным совпадением. Кроме того, удержание клавиши управления временно превращало центральный диск в серый до тех пор, пока клавиша не была отпущена. Участников попросили стереть нежелательные послеобразы от настроенного центрального диска, используя эту клавишу, прежде чем подтвердить настройку. Таким образом, они удерживали клавишу управления, ждали и двигали глазами, пока центральный диск не становился ахроматическим. Затем они смотрели на серый круг до тех пор, пока движущийся круг не достиг максимальной насыщенности. Только после этого они отпускали клавишу и сравнивали ранее настроенный цвет с цветом движущегося круга. Как правило, настроенный цвет был слишком насыщенным из-за наложения его собственного послеобраза во время настройки. Поэтому участникам пришлось снизить насыщенность после повторной адаптации. Они повторяли эту процедуру до тех пор, пока насыщенность не требовала дальнейшей регулировки после отпускания клавиши управления. Только тогда участники подтверждали свои настройки, нажимая Enter. Как и в Эксперименте 1, использовался межпробный период для предотвращения переноса послеобразов между испытаниями.
Индукторные цвета были разделены на девять серий по восемь цветов. В пределах серии восемь цветов разделялись 45 градусами по азимуту. Различные серии определялись разными стартовыми точками (0, 5, 10…40 градусов), так что серии вместе охватывали все 72 индуктора. Каждый блок измерений включал одно испытание для каждого из восьми индукторных цветов в серии, представленных в случайном порядке. Измерения каждого блока повторялись до пяти раз (см. рис. S8–10). Блоки измерений были распределены на несколько дней. Перед измерениями участники проходили обучение с пробными блоками под наблюдением и с обратной связью от экспериментатора (CW), чтобы убедиться, что они поняли обработку задачи и цель максимально точного измерения цвета иллюзорного послеобраза.
Модели
Во всех моделях цветовой сигнал серого пробного круга (того же, что и фоновый цвет) рассчитывался при локальной адаптации к индуктору. Затем цвет результирующего послеобраза определялся как локально адаптированный цветовой сигнал при глобальной адаптации к серому фону. Известный факт, что послеобразы не так насыщены, как индукторы (Рис. 2f), подразумевает, что адаптация к индукторам не является полной. В качестве оценки силы адаптации, адаптирующая насыщенность для моделирования послеобразов была установлена на уровне насыщенности сравнительных цветов в Эксперименте 1 и на уровне общей средней насыщенности (27.0 в CIELUV и 0.47 в DKL) настроек в Эксперименте 2. Даже если бы предполагалась полная адаптация к насыщенности индукторов, результаты были бы в значительной степени теми же. Однако, неудивительно, коэффициенты корреляции, включающие адаптацию колбочек, были бы немного ниже, поскольку нелинейности в модели в этом случае выше, чем в измерениях. В моделях нет свободных параметров.
Адаптация первого этапа
Адаптация колбочек на первом этапе цветовой обработки моделировалась с помощью контрастов колбочек. Контрасты колбочек (CC) – это доли Вебера, рассчитанные как разница между возбуждениями колбочек стимула (LMS) и возбуждениями колбочек адаптирующего цвета (LMS0), отнесенная к возбуждениям колбочек адаптирующего цвета: CC = (LMS - LMS0) / LMS0. Контраст колбочек рассчитывается независимо для коротковолновых (S), средневолновых (M) и длинноволновых (L) чувствительных колбочек, что приводит к контрастам S-колбочек, M-колбочек и L-колбочек. В психофизических экспериментах адаптация обычно контролируется цветом фона, который затем является LMS0. Для вычисления индуцированного цвета послеобраза роли меняются, поскольку послеобразы соответствуют восприятию ахроматического фона после локальной адаптации к индуктору. Локальная адаптация к индуктору моделируется путем вставки возбуждений колбочек индуктора, iLMS, вместо фона в LMS0. Поскольку послеобраз вызывается на сером пробнике, LMS теперь соответствует bgLMS, т.е. ахроматическому, изолюминантному серому цвету фона:
(1) Адаптация колбочек к индуктору:
$$CC_{LMS}= rac{bgLMS - iLMS}{iLMS}$$
Где bgLMS и iLMS относятся к возбуждениям колбочек по Стокману-Шарпу ахроматического фона и адаптирующего индуктора. Возбуждения колбочек масштабировались для соответствия функции световой эффективности. Контраст колбочек изменяется с увеличением адаптации колбочек iLMS в соответствии с обратно пропорциональной функцией (поскольку iLMS увеличивается в знаменателе). Эта адаптация производит смещения к пиковым чувствительностям колбочек (Рис. 1b, Рис. S14).
Адаптация второго этапа
Колбочная оппонентность моделировалась с использованием цветового пространства DKL. Подробности вычисления колбочно-оппонентных осей приведены в разделе E.2 Модель колбочно-оппонентности (DKL) Дополнительных материалов. Предполагается, что адаптация колбочно-оппонентности является субтрактивной. Поэтому она рассчитывалась путем вычитания колбочно-оппонентного сигнала адаптирующего цвета из колбочно-оппонентного сигнала ахроматического пробника.
(2) Адаптация колбочно-оппонентности к индуктору:
$$DKL_{LM} = bgDKL_{LM} - k * iDKL_{LM}$$
$$DKL_S = bgDKL_S - k * iDKL_S$$
Где DKLL-M и DKLS – адаптированные колбочно-оппонентные сигналы механизмов L-M и S соответственно, а bgDKL и iDKL – колбочно-оппонентный сигнал фона и индуктора соответственно. Постоянная k указывает силу адаптации. Поскольку фон ахроматичен, bgDKL равен 0 во всех каналах, и уравнения могут быть упрощены до:
$$DKL_{LM} = -k * iDKL_{LM}$$
$$DKL_S = -k * iDKL_S$$
Следовательно, адаптация колбочно-оппонентности производит пропорциональные сдвиги вдоль осей DKL, предсказывая, что оттенки колбочно-оппонентных послеобразов будут повернуты на 180 градусов от индуктора, а насыщенность послеобразов будет k раз больше насыщенности индуктора. Здесь k было установлено для соответствия оцененной силе послеобразов, но значение k не влияло на результаты. При изолюминансности колбочно-оппонентные механизмы могут быть выражены как функция контрастов колбочек, что приводит к:
$$DKL_{LM} = (iCC_M - iCC_L)$$
$$DKL_S = iCC_S$$
Где iCC – контраст колбочек индукторов, рассчитанный по уравнению (1), а индекс указывает соответствующую колбочку (см. раздел E.2 Модель колбочно-оппонентности (DKL) Дополнительных материалов). Хотя теоретически полная модель различала бы два отдельных механизма, составляющих каждый оппонентный канал («половинная ректификация»), такое разделение не изменило предсказаний в этом исследовании.
Адаптация цветового зрения
Предсказания моделей цветового зрения (CIELUV, CIELAB, CIECAM02) вычислялись как цветовое зрение ахроматического фона при условии, что адаптирующая точка белого является индуктором, с использованием соответствующего преобразования адаптации. При изолюминансности CIELUV является проекционным преобразованием пространства DKL и включает субтрактивную адаптацию, что объясняет идентичность предсказываемых оттенков. Разделы E3-E5 Дополнительных материалов содержат математические детали и объяснения. Для предсказаний цветов Манселла и Херинга оппонентные цвета определялись в соответствующих координатных системах, что эквивалентно эффектам субтрактивной адаптации. Оппонентные цвета в системе Манселла интерполировались с использованием координат CIELAB из таблицы перенумерации Манселла, предоставленной Лабораторией Манселла Рочестерского технологического института. Подробности интерполяций приведены в разделе E.6 Модель оппонентности Манселла Дополнительных материалов. Херенговские оппонентные цвета рассчитывались путем линейной интерполяции направления оттенка между соответствующими двумя эмпирически измеренными прототипами для красного, желтого, зеленого и синего в пространстве CIELUV. Предсказаний для насыщенности Херинга не было. Подробности см. в разделе E.7 Модель оппонентности Херинга Дополнительных материалов.
Распределения данных
Измерения оттенка и насыщенности были приблизительно нормально распределены по повторным измерениям и участникам, как было проверено гистограммами (Рис. S3–S5) и графиками нормального распределения (Рис. S6–S7).
Результаты: Ключевые выводы исследования
На рисунке 1 представлены цветовые стимулы (Рис. 1a) и соответствующие предсказания оттенка и насыщенности послеобразов кандидатными моделями послеобразов (Рис. 1b–e). Предсказания для адаптации колбочек были рассчитаны с использованием математической модели закона Вебера. Использование индуктора в качестве адаптирующего цвета приводит к треугольной форме красной кривой на Рис. 1b. Треугольная форма возникает из-за того, что предсказываемый цвет доминируется наименее адаптированной колбочкой из-за обратно пропорциональных эффектов адаптации колбочек (см. Метод и Рис. S14). Эта треугольная форма означает, что адаптация колбочек противоречит распространенному предположению, что послеобразы являются колбочно-оппонентными.
В отличие от этого, колбочно-оппонентные послеобразы возникают в результате субтрактивной, вторичной адаптации, как показано на красном круге на Рис. 1c. Были предложены альтернативные модели, чтобы лучше предсказывать несколько аспектов цветового зрения, чем эти физиологические модели. Они могут лучше соответствовать и цветовому зрению послеобразов. CIELUV, CIELAB и CIECAM02 реализуют сложные, нелинейные модели для аппроксимации различных эмпирических данных о цветовом зрении. При изолюминансности (как здесь) CIELUV предсказывает те же оттенки послеобразов, что и колбочная оппонентность, но другие насыщенности послеобразов (Рис. 1c; см. Метод). CIELAB и CIECAM02 (Рис. 1d) включают преобразования адаптации, вдохновленные адаптацией фон Криса, и производят треугольную форму, похожую, но иначе ориентированную, чем у адаптации колбочек. Система Манселла и цвета Херинга (Рис. 1e) предлагают еще один подход к идентификации комплементарных цветов, которые считались точками отсчета для цветового зрения. Эксперименты 1 и 2 проверили предсказания с варьирующимся оттенком индуктора, используя две принципиально разные задачи. Эксперимент 3 был посвящен эффекту варьирующейся насыщенности индуктора на оттенок послеобраза. Все данные и код доступны через Zenodo. Код для моделей адаптации колбочек и колбочной оппонентности также включен и объяснен в разделах E.1–2 Дополнительных материалов.
Эксперимент 1: Сравнение цветовых послеобразов
Первая задача сочетала типичный дисплей послеобразов с одновременным совпадением (Рис. 2a). Наблюдатели сопоставляли оттенок сравнительных стимулов с оттенком послеобраза, который появляется в месте индукторного цвета после исчезновения индукторного цвета. Насыщенность сравнительных стимулов оставалась постоянной (см. Метод).
В первой части (Эксперимент 1a) измерения с высоконасыщенными цветами показали, что оттенки послеобразов не были ни колбочно-оппонентными, ни херенговско-оппонентными к оттенкам индукторов (Рис. 2b). Оттенки послеобразов для шести из восьми индукторов значительно отличались от колбочно-оппонентных оттенков (Таблица 1; см. также рис. S2a, S3 и Таблица S2). В задачах именования оттенки послеобразов красного, желтого, зеленого и синего назывались синими, пурпурными, пурпурно-розовыми и оранжево-коричневыми, что резко контрастирует с красно-зеленой и сине-желтой оппонентностью Херинга (см. Таблицу 1; подробности см. в Таблицах S2–S3, Рис. S3). Важнее всего то, что измеренные оттенки послеобразов были близки к предсказаниям адаптации колбочек (Рис. 2i и Рис. S2), а их отклонения от колбочно-оппонентных оттенков почти идеально коррелировали с отклонениями, предсказанными адаптацией колбочек (Пирсон r(6) = 0.98, 95% ДИ = [0.88, 1.00], p).
Эксперимент 2: Динамическое предъявление стимулов
Во втором эксперименте использовалась «гоняющаяся» парадигма, где индукторный цвет постоянно перемещался по кругу, а наблюдатель должен был найти соответствующий послеобраз. Этот метод позволил точно измерить послеобразы, выявить их основные характеристики и сравнить с предсказаниями моделей. Результаты подтвердили, что послеобразы следуют предсказаниям адаптации колбочек, а не колбочной оппонентности. Это наблюдалось как в ответах на оттенок, так и на насыщенность.
В частности, было обнаружено, что адаптация колбочек может объяснить более 60% вариативности (дисперсии) в восприятии оттенка и более 80% вариативности в восприятии насыщенности послеобразов (Рис. 2). Несмотря на различия между участниками, все индивидуальные данные предоставили недвусмысленные доказательства сигнатуры адаптации колбочек при учете индивидуальных различий в силе индукции в модели (Рис. S8–S10). Изменения оттенка послеобразов по насыщенности также следовали модели адаптации колбочек (Рис. 3). Учитывая шум измерения, эти результаты предполагают, что комплементарные цвета в послеобразах полностью определяются адаптацией колбочек на первом этапе цветовой обработки.
Модель адаптации колбочек: Детальное объяснение
Используемая здесь модель адаптации колбочек реализует дивергентную адаптацию в соответствии с законом Вебера, что означает, что чувствительность колбочек снижается пропорционально величине текущего возбуждения колбочек. Согласно классическому объяснению послеобразов через «выцветание колбочек», эта модель отражает пропорциональное снижение количества уловленных фотонов фоторецепторами. Данный свет имеет определенную вероятность, с которой он изомеризует молекулы ретиналя, связанные с опсином. Когда молекулы ретиналя уже претерпели изомеризацию из-за текущей стимуляции индуктором, эта вероятность применяется только к оставшейся доле молекул ретиналя, что подразумевает, что количество изомеризующихся молекул ретиналя пропорционально доступным, неизомеризованным молекулам. В результате адаптированный отклик колбочек пропорционален обратной величине изомеризованных индуктором молекул. Образование послеобразов можно объяснить временным лагом между изомеризацией колбочек и регенерацией.
Классические критики объяснения выцветания колбочек включали различия между послеобразами от чередующихся и статических индукторов, продолжительность послеобразов (>10 с) намного дольше, чем адаптация колбочек («100 мс), и низкий уровень изомеризации при обычной, умеренной яркости. Эти критики кажутся сомнительными. Во-первых, схожие хроматические послеобразы для чередующихся и статических индукторов были обнаружены недавно, а восприятие послеобразов от динамических индукторов может зависеть от факторов, не связанных с адаптацией рецепторов. Во-вторых, было признано, что регенерация колбочек может занимать минуты, а не миллисекунды. В-третьих, уровни изомеризации соответствуют откликам колбочек адаптирующего стимула, здесь – индуктора. Кажется противоречивым, что уровень изомеризации при обычной яркости (1–10%) достаточен для создания насыщенных неиллюзорных цветов индукторов, но считается недостаточным для объяснения менее насыщенных послеобразов.
В то же время стало ясно, что адаптация колбочек при умеренных уровнях освещенности включает процессы, отличные от «выцветания колбочек». Кроме того, временная динамика адаптации сложна и различается для разных типов колбочек. Задачи в настоящем исследовании были разработаны для измерения цветов послеобразов при максимальной адаптации и для избежания временных эффектов во время формирования и затухания послеобразов. Несмотря на сложность лежащих в основе физиологических процессов, результирующая дивергентная адаптация колбочек к умеренно ярким, фотопическим цветам может быть приближена контрастами колбочек в соответствии с законом Вебера.
Адаптация на уровне колбочек и ее роль
После адаптации колбочек адаптированный сигнал колбочек распространяется через последующие стадии обработки цвета. Сигнал отскока, наблюдаемый в ганглиозных клетках сетчатки, указывает на то, что адаптированный сигнал колбочек временно поддерживается на второй, колбочно-оппонентной стадии. Это временное поддержание, вероятно, способствует устойчивой продолжительности послеобразов. Согласно настоящим результатам, обработка цветового сигнала на второй стадии, по-видимому, не влияет на воспринимаемые цвета послеобразов каким-либо специфическим образом.
Вместо этого дивергентная, подобная Веберу адаптация колбочек объясняет, почему оттенки послеобразов не являются колбочно-оппонентными, не взаимными и производят три кластера оттенков. Обратно пропорциональная функция возбуждения колбочек индуктора создает нелинейные эффекты адаптации колбочек, которые приводят к трем кластерам оттенков и трем пикам насыщенности послеобразов по сравнению с колбочной оппонентностью (Рис. 2d, f, h). Каждый из трех пиков послеобразов определяется наименее адаптированной, а не наиболее адаптированной колбочкой (см. Рис. S14): Если L- и M-колбочки наиболее адаптированы индукторами, S-колбочки доминируют в послеобразе; если L- и S-колбочки адаптированы, M-колбочки доминируют в послеобразах; и если S- и M-колбочки наиболее адаптированы, L-колбочки доминируют в послеобразах. Чувствительность L- и M-колбочек перекрывается. Таким образом, максимальная адаптация одной всегда подразумевает некоторую адаптацию другой, и три локальных максимума (пика) модели адаптации колбочек (красная кривая на Рис. 1b) не совпадают с направлениями изолированных возбуждений колбочек (серые линии на Рис. 1b). Перекрытие между L- и M-колбочками сильнее для широкополосных спектров ненасыщенных цветов, что приводит к смещению оттенка от изолированных возбуждений колбочек с уменьшением насыщенности (изогнутые черные линии на Рис. 3b). Поскольку чувствительность S-колбочек сравнительно мало перекрывается с чувствительностью M- и L-колбочек, эффект S-колбочек на L-M-адаптированные послеобразы намного сильнее, чем эффекты коррелированных L- или M-колбочек, что приводит к выраженной асимметрии вдоль колбочно-оппонентной оси S-(L+M) (близко к сине-желтой оси v*, см. Рис. 1b, 3c и S11.b).
Неоппонентность: опровержение старых теорий
Напротив, субтрактивная адаптация колбочно-оппонентного механизма не способна предсказать три кластера оттенков и асимметрию вдоль механизма S-(L+M) (Рис. 1c и S11.c). Неспособность предсказать асимметрию объясняет, почему субтрактивная адаптация в пространстве DKL относительно неуспешна в предсказании насыщенности послеобразов (Рис. 2j). Поверхностный взгляд на послеобразы может вводить в заблуждение, поскольку послеобразы на грубом уровне напоминают колбочную оппонентность. Это происходит не из-за адаптации колбочно-оппонентных механизмов на второй стадии цветовой обработки, а из-за того, что колбочно-адаптированный цветовой сигнал с первой стадии распространяется на вторую и последующие стадии. Процедуры обнуления послеобразов (настройки, которые отменяют послеобраз) в колбочно-оппонентном пространстве также не позволяют продемонстрировать неоппонентность послеобразов.
Успешное обнуление работает независимо от того, следует ли появление послеобразов по прямой линии из-за субтрактивной колбочно-оппонентной адаптации или по кривой из-за нелинейной, дивергентной адаптации, такой как адаптация колбочек. Хотя отклонения оттенков послеобразов от колбочной оппонентности тонки и требуют специально разработанных методов для эмпирического измерения, они достаточно сильны, чтобы быть видимыми при личном восприятии. Это можно продемонстрировать, используя «гоняющуюся» схему на стандартном компьютерном дисплее, как в приложенных анимированных визуализациях (Дополнительные фильмы 1–4; инструкции см. в разделе A Дополнительных материалов).
CIELAB и CIECAM02 дают лучшие предсказания, чем колбочная оппонентность, поскольку их псевдо-фон-крисовские трансформации адаптации производят 3 кластера, близких к адаптации колбочек, и улавливают часть асимметрии вдоль механизма S-(L+M). Тем не менее, они отличаются от адаптации колбочек тем, что CIELAB и CIECAM02 используют псевдо-фон-крисовское преобразование, применяемое к первичным цветам, которые похожи, но не идентичны колбочкам (см. дополнительные разделы E.4–5 для подробностей). Это предполагает, что включение надлежащего преобразования адаптации колбочек в модели цветового зрения улучшило бы их способность предсказывать комплементарные послеобразы.
Несмотря на то, что системы Манселла и херенговская оппонентность долгое время были фаворитами для объяснения явлений цветового зрения, они являются худшими предсказателями цветов послеобразов (Рис. 2i, j). Они предсказывают, что послеобразы смещаются от колбочной оппонентности (Рис. 1d) в направлениях, противоположных предсказаниям адаптации колбочек (Рис. 1b). Различия между послеобразами и херенговской оппонентностью настолько велики, что соответствующие цветовые термины – красный, желтый, зеленый и синий – даже не являются грубыми приближениями комплементарности послеобразов (Рис. S3, Таблица 1, S2–S3). В совокупности эти выводы опровергают самое распространенное предположение, что комплементарные послеобразы являются цветооппонентными в какой-либо из широко распространенных интерпретаций.
Кортикальная адаптация: роль и ограничения
Другие утверждали, что цвета послеобразов формируются через адаптацию гипотетических кортикальных механизмов, включающих распознавание формы и объектов, бинокулярную интеграцию и/или цветовую постоянство, что противоречит классическим доказательствам ретинального происхождения. Эффекты «сверху вниз» на цветовое зрение хорошо известны. Такие эффекты могут быть особенно сильными для иллюзорных перцептов, таких как послеобразы, где физический стимул отсутствует. Однако доказательства эффектов среднего уровня (например, контуров и форм) или высокого уровня (например, знаний) на послеобразы не обязательно отражают происхождение послеобразов. Послеобразы возникают в отсутствие стимулов или знаний «сверху вниз», как в настоящих экспериментах. Следовательно, помехи «сверху вниз» могут модулировать субъективное восприятие послеобразов; но они не являются причиной послеобразов.
Модель адаптации колбочек оставляет мало необъясненной дисперсии воспринимаемых послеобразов (оценочно 20% с сглаженными данными), которая потребовала бы дополнительной адаптации на более поздних, последующих уровнях обработки. Большая часть этой дисперсии, скорее всего, является результатом шума и индивидуальных различий (см. Ограничения ниже). Поэтому спекуляции о еще неизвестных кортикальных механизмах для объяснения комплементарных цветов послеобразов не являются необходимыми.
Ограничения исследования
Несмотря на индивидуальные различия в чувствительности колбочек, настоящий подход предполагал одинаковую чувствительность колбочек стандартного наблюдателя CIE для всех индивидуумов. Модель адаптации колбочек, основанная на индивидуальной чувствительности колбочек, могла бы дать еще лучшие предсказания и объяснить часть индивидуальных различий, наблюдаемых в измерениях послеобразов (Рис. S8–S10).
Несмотря на достоверность модели адаптации колбочек, она не информирует нас о точных физиологических процессах, которые реализуют эти Вебер-подобные вычисления. Следует также отметить, что адаптация колбочек, проявляющаяся в послеобразах, является важным, но не единственным фактором, способствующим адаптации цвета и постоянству в естественных условиях просмотра. Стимульные дисплеи в этом исследовании были разработаны специально для измерения цветов послеобразов при контроле других источников цветовой индукции, таких как индукция локального контраста, распознавание форм, иллюзорные контуры/области или осмысленные объекты. Цвета, воспринимаемые при адаптации к более сложным конфигурациям стимулов, включая естественные сцены и другие реальные среды, вероятно, определяются комбинацией различных механизмов адаптации и когнитивных эффектов «сверху вниз».
Заключение: Переосмысление цветовых послеобразов
Таким образом, настоящие результаты показывают, что количественная модель адаптации колбочек достаточна для объяснения перцептивной природы комплементарных цветов в послеобразах. Это понимание разрешает противоречия между перцептивной природой послеобразов и теориями об их физиологическом происхождении, которые были предметом многолетних дебатов. В отличие от того, что предполагают современные учебники, цветовые послеобразы не являются результатом колбочной или херенговской цветовой оппонентности. Вместо этого они могут быть полностью и последовательно объяснены хорошо обоснованной, прямолинейной моделью адаптации колбочек на первом этапе цветовой обработки.
Кроме того, хроматическая адаптация, выявляемая послеобразами, играет фундаментальную роль в зрительном восприятии, поскольку она также модулирует чувствительность к цвету, обеспечивает цветовую постоянство и определяет, как цвета субъективно выглядят для наблюдателей. Например, дивергентная адаптация колбочек позволяет нам воспринимать цвета постоянно при изменениях освещения, компенсируя дивергентные цветовые сдвиги (улавливаемые стабильными соотношениями возбуждения колбочек), которые типичны для изменений освещения в естественных условиях. Как показано на этих широко используемых моделях цветового зрения (Рис. 1b, c), лучшее понимание эффектов адаптации на цветовое зрение также является ключом к измерению, спецификации и контролю цветов (цветовые измерения) в научных и промышленных применениях.