Интересное сегодня
Мир полон неопределенностей и опасностей. На протяжении всей эволюционной истории большинства животных, несмотря на снижение уровня смертности для современного человека, мы сталкиваемся с различными рисками. Восприятие человеком риска не всегда коррелирует с его объективной вероятностью. Например, ядерная энергетика, считающаяся экспертами относительно безопасной, часто воспринимается неспециалистами как чрезвычайно рискованная. Подобное искаженное восприятие может привести к экономическим издержкам, например, в случае перерасхода средств на борьбу с терроризмом в ущерб другим насущным потребностям, таким как образование или здравоохранение.
Понимание страха перед катастрофами
Общеизвестно, что люди склонны избегать ситуаций, воспринимаемых как недобровольные, неконтролируемые, смертельно опасные или несправедливые, особенно когда они затрагивают большое количество людей одновременно. Такие риски называют "рисками страха" (dread risks). Чем более неизвестными, незнакомыми и системными являются причины таких опасностей, тем сильнее люди склонны считать, что такие обстоятельства предсказывают будущие риски.
Терроризм как тактика повстанцев, похоже, эксплуатирует психологические особенности "страха". Влиятельный анализ поведения американцев после терактов 11 сентября 2001 года демонстрирует, насколько сильной может быть реакция "страха" на терроризм. Существуют убедительные доказательства того, что путешествие на автомобиле гораздо опаснее, чем на самолете. Анализ, проведенный Гердом Гигеренцером, показал, что после 11 сентября американцы стали избегать авиаперелетов, предпочитая автомобильные поездки, что привело к примерно 1595 дополнительным случаям гибели людей на дорогах. Эти случаи можно рассматривать как "косвенный ущерб от терроризма", вызванный реакцией "страха" на атаки. Гигеренцер интерпретировал эту реакцию как неадаптивную, обусловленную неспособностью точно оценить относительные риски различных видов транспорта, и призвал к информированию людей об этих рисках в рамках психологически обоснованной "расширенной контртеррористической политики".
Хотя в текущих условиях предпочтение автомобиля может показаться нерациональным, мы покажем, как такая реакция, даже при полном информировании об относительных рисках, может быть адаптивной в определенной среде, если правильно учитывать эволюционные последствия смертельных рисков различных масштавов.
Эволюционные корни страха: теория
Теоретических работ, объясняющих эволюционное происхождение реакций "страха", существует немного. Наиболее проработанные идеи фокусируются на адаптивной ценности реакций "страха" на воспринимаемое увеличение рисков массовой гибели. Было предложено, что такое поведение, задокументированное после 11 сентября, могло быть отобрано у наших предков. Например, в работе Бодемера и соавторов (Bodemer et al.) "страх" анализируется через призму влияния событий, приводящих к гибели, на показатель совокупной численности популяции. Мы согласны с Бодемером и соавторами в том, что организмы должны эволюционировать таким образом, чтобы гибель от одних источников считалась хуже, чем от других. Однако их модель не включает генетический компонент, упуская тем самым суть проблемы.
Естественный отбор действует на частоту генов, и с этой точки зрения индивиды важны лишь постольку, поскольку они передают гены будущим поколениям. Одни источники смертности затрагивают носителей определенного гена независимо друг от друга (демографический или идиосинкратический риск), другие же уничтожают значительную долю всех носителей гена, если происходят (экологический или совокупный риск). В настоящей работе мы формально покажем, как могут эволюционировать существенные предубеждения против последних, даже когда они подвергают индивидов более высокому уровню демографического риска, и таким образом объясним реакции "страха", подобные тем, что связаны с террористическими атаками, приводящими к массовой гибели.
Детали модели
Мы предполагаем, что рискованное поведение индивида является унаследованным признаком с генетической основой. Наша цель — предсказать значение этого признака, которое будет эволюционировать под действием естественного отбора. Для этого мы используем стандартный подход и предполагаем, что признак определяется аллелем в одном аутосомном локусе, признавая при этом, что в реальности рискованное поведение, вероятно, находится под влиянием множества генетических локусов. При нашем упрощающем допущении различные аллели в данном локусе определяют различное рискованное поведение. Поведение, определяемое аллелем, влияет на количество потомства, оставляемого носителями аллеля, и, следовательно, на количество копий аллеля в следующем поколении. Таким образом, количество копий аллеля в популяции изменяется с характерной скоростью по поколениям. Эта скорость изменения известна как инвазивная приспособленность аллеля. Затем ожидается, что действие естественного отбора приведет к популяции, в которой большинство индивидов несут аллель с наибольшей инвазивной приспособленностью. Таким образом, при эволюционной стабильности мы ожидаем, что эволюционировавшее рискованное поведение будет максимизировать инвазивную приспособленность аллеля, ответственного за это поведение.
Когда мы фокусируемся на конкретном аллеле, мы делаем упрощающее допущение, что все носители аллеля имеют только одну его копию. Таким образом, размер когорты индивидов, несущих аллель, равен общему количеству копий аллеля. Скорость увеличения числа аллелей (инвазивная приспособленность) тогда равна скорости увеличения числа когорт. Эта скорость увеличения зависит от воспроизводства и смертности. Все индивиды размножаются непрерывно: скорость передачи фокального аллеля потомству равна R, независимо от влияния аллеля на рискованное поведение. Индивиды подвергаются двум источникам смертности, уровень которой зависит от рискованного поведения и, следовательно, от интересующего нас аллеля. Один источник смертности — демографический, то есть различные члены когорты затрагиваются независимо (идиосинкратический риск). Смертность на одного индивида от этого риска составляет M_D. Другой источник — экологический (совокупный риск). Он моделируется путем предположения о периодических экологических катастрофах. Эти катастрофы происходят по пуассоновскому процессу с интенсивностью ambda. Когда происходит катастрофа, умирает фиксированная доля p когорты. Таким образом, смертность от катастроф на одного индивида, несущего фокальный аллель, составляет M_C = ambda p.
В приведенном выше уравнении ambda является фиксированным экологическим параметром. В отличие от него, параметры M_D и p зависят от рискованного поведения и, следовательно, от фокального аллеля.
Результаты: инфляция рисков
Мы делим наши результаты на две части. Сначала мы выводим формулу для инвазивной приспособленности аллеля, учитывающую два источника смертности, связанных с аллелем. Эта формула подчеркивает, что два источника смертности не эквивалентны. Во второй части мы предполагаем, что индивиды сталкиваются с компромиссом между двумя источниками смертности и анализируем, как, по нашим ожиданиям, естественный отбор будет формировать отношение к риску в условиях этого компромисса.
Скорость увеличения числа аллелей и инфляция
При расчете инвазивной приспособленности аллеля мы должны учитывать два источника стохастичности. Идиосинкратический риск действует независимо на разных носителей аллеля, поэтому мы можем просто усреднить этот источник стохастичности, когда число носителей аллеля велико. Напротив, совокупный риск затрагивает многих носителей одинаково, поэтому такая простая форма усреднения неуместна. Это хорошо известно в азартных играх и инвестировании, когда цель — максимизировать долгосрочный рост капитала. В этих сценариях, даже если ставка или инвестиция имеет самую высокую среднюю доходность, доступную в настоящее время, если ставка рискованна, лучше направить в нее только часть текущего капитала на случай невезения. Здесь критерий Келли (Kelly criterion) предоставляет стратегию управления деньгами, которая максимизирует геометрическую скорость роста капитала. Следует отметить, что максимизация геометрической скорости роста эквивалентна максимизации ее логарифма. При рассмотрении инвазивной приспособленности долгосрочное увеличение числа аллелей аналогично долгосрочному росту капитала. Как показано в Методах (ниже), следует усреднять логарифмическое увеличение числа аллелей как меру инвазивной приспособленности.
В Методах мы показываем, что инвазивная приспособленность определяется формулой:
$$r = R (M_D + lpha M_C)$$
где:
$$ lpha = rac{og (1-p)}{p} $$
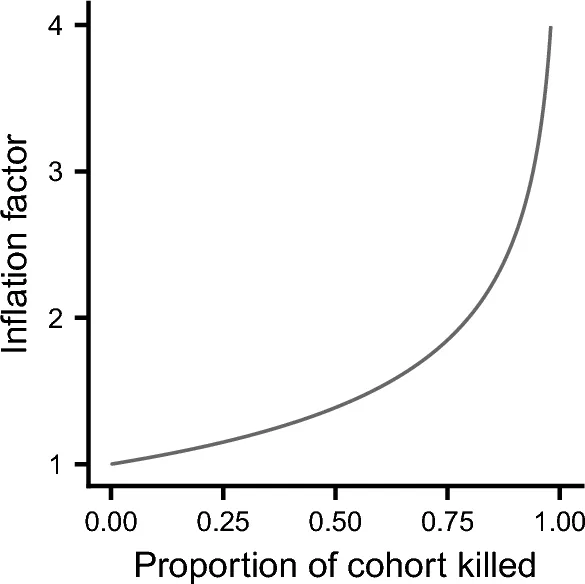
Эти формулы демонстрируют, что два источника смертности не эквивалентны по своему влиянию на инвазивную приспособленность. В частности, риск от экологического источника смертности умножается на фактор lpha по сравнению с риском от демографического источника смертности. Фактор инфляции всегда больше 1 и увеличивается по мере роста доли p когорты, погибающей в катастрофе. Рисунок 1 показывает зависимость фактора инфляции от p. Так, например, если 60% когорты погибает в катастрофе, то влияние этого источника смертности на инвазивную приспособленность увеличивается примерно на 50%. Чтобы проиллюстрировать это, предположим, что катастрофы происходят в среднем каждые 20 лет и уносят жизни 60% когорты. Тогда фактическая смертность от катастроф составит 1 смерть на душу населения каждые 30 лет. Однако катастрофы снизят инвазивную приспособленность примерно так же, как демографический источник смертности, вызывающий одну смерть на душу населения каждые 20 лет.
Распределение риска
Теперь предположим, что члены популяции могут выбирать, какому источнику смертности подвергаться. Представим это как индивидуальные поездки на автомобиле против групповых поездок на самолете. Мы предполагаем, что члены популяции должны совершать относительно безопасное путешествие каждый день или неделю. Если они используют автомобиль, то погибают в пути с вероятностью u, независимо от других членов популяции. Если они используют самолет, то самолет терпит крушение с вероятностью ambda, и все индивиды, которые в этот день летят на самолете, погибают. Для простоты мы предполагаем, что нет зависимости от частоты, влияющей на эти уровни смертности. В частности, количество членов популяции, выбирающих автомобиль, и, следовательно, плотность автомобилей на дорогах, не влияет на вероятность u того, что данный автомобиль потерпит крушение. Аналогично, количество людей, решающих лететь на самолете, не влияет на то, произойдет ли крушение конкретного самолета.
Мы предполагаем, что решение о форме транспорта определяется аллелем, который определяет вероятность p того, что носитель аллеля выбирает самолет при каждой поездке. Выбор автомобиля или самолета одним индивидом независим от выбора других. Как и прежде, нас интересует когорта индивидов, несущих этот аллель, и мы рассматриваем скорость роста численности когорты. При предположении о достаточно большом числе индивидов (чтобы мы могли усреднить демографическую стохастичность), доля членов когорты, использующих самолет при каждой поездке, равна p, следовательно, M_C = ambda p и M_D = (1-p)u. Согласно Уравнениям (1) и (2), инвазивная приспособленность аллеля равна:
$$r(p) = R(1-p)u + ambda og (1-p)$$
Пусть p^* — значение p, максимизирующее эту меру приспособленности. Поскольку нет зависимости от частоты, аллель, кодирующий эту вероятность полетов на самолете, будет распространяться до фиксации в любой резидентной популяции. Таким образом, полеты на самолете с этой вероятностью являются уникальной эволюционно стабильной стратегией.
Дифференцируя по p, мы получаем:
$$ rac{dr}{dp} = u - rac{ambda }{1-p} $$
Таким образом, если ambda > u, инвазивная приспособленность всегда уменьшается с увеличением p, следовательно, p^*=0; то есть при эволюционной стабильности все едут на машине. Однако, если ambda < u, при эволюционной стабильности доля:
$$ p^*=1 - rac{ambda }{u } $$
летит на самолете. Обратите внимание, что в этом случае фактическая смертность на душу населения от использования автомобиля составляет (1-p^*)u = ambda, в то время как от использования самолета — p^*ambda. Таким образом, независимо от того, насколько самолет безопаснее автомобиля, больше членов когорты погибает в автомобильных авариях, чем в авиакатастрофах. В пределе, когда самолет становится сверхбезопасным по сравнению с автомобилем (ambda l u, так что p^* pprox 1), количество погибших в авиакатастрофах и автомобильных авариях почти равно.
Обсуждение
Наш подход предполагает, что отношение к риску имеет генетическую основу, причем аллели в аутосомном локусе определяют отношение носителя аллеля. Стандартным аргументом является то, что в точке эволюционного равновесия аллель, который был отобран, должен максимизировать свою инвазивную приспособленность. Это означает, что скорость роста числа аллелей максимизируется. Соответствующей мерой "роста" является геометрическое среднее увеличение числа копий, пока аллель редок, или обобщение этой концепции для структурированных популяций. Риски варьируются по степени корреляции их влияния на разных индивидов, несущих аллель, между двумя крайностями. На одном крайнем уровне, при демографическом (идиосинкратическом) риске, таком как невезение в поиске пищи, гибель индивида, несущего аллель, независима от гибели других носителей. На другом крайнем уровне, при экологическом (совокупном) риске, таком как плохая погода, значительная доля всех носителей аллеля погибает.
Давно известно, что форма риска имеет решающее значение для измерения инвазивной приспособленности. Кроме того, эта работа подчеркивает, что экологический риск оказывает непропорциональное влияние по сравнению с демографическим риском. Например, Робсон (Robson) отмечает, что критерий приспособленности "включает существенное различие между идиосинкратическим риском в данных условиях и совокупной неопределенностью в отношении самой среды, подразумевая большее неприятие последнего. Это связано с тем, что текущая модель порождает форму автоматического биологического страхования посредством закона больших чисел против идиосинкратического риска, тогда как это страхование не действует в том же смысле против совокупной неопределенности". Этот эффект проиллюстрирован в контексте финансовых инвестиций.
Эти аргументы можно переформулировать, чтобы сделать вывод, что если эволюция приводит к тому, что индивид безразличен к демографическому и экологическому риску, то вероятность гибели при экологическом риске должна быть меньше. Наш вклад в эту работу заключается в точном количественном определении, насколько меньше, причем фактор инфляции, связывающий вероятности гибели при двух формах риска, зависит только от доли родственников, погибающих в случае события, связанного с совокупным риском.
Когда возникает новый аллель, то, пока он еще редок, если индивид несет аллель, другие его носители являются родственниками. Таким образом, эффект инфляции должен наблюдаться при ограниченном потоке генов между группами ранних людей. Ограниченный поток генов означает, что почти все копии аллеля находятся в группе, где он возник, поэтому риск, затрагивающий всю группу, является совокупным риском. Напротив, при значительном потоке генов риск, затрагивающий только одну группу, действует скорее как демографический риск. Этот контраст проиллюстрирован в работе [23], где наблюдается отказ от рискованного поведения по мере увеличения скорости расселения между локальными группами. Данные современных охотников-собирателей предполагают значительное перемещение индивидов между группами, поэтому, на первый взгляд, ранние люди могли не испытывать ограниченного потока генов. Даже если бы ранние люди имели ту же социальную структуру, что и современные охотники-собиратели, это не является убедительным, поскольку анализ в [23] предполагает, что опыт каждой группы независим. Если группы не независимы (например, потому что они испытывают одинаковые погодные условия), расселение не устранит совокупный риск, поскольку носители фокального гена в разных группах будут испытывать схожие условия.
В современном мире многие предпочитают ездить на машине, а не летать. Такое избегание самолетов не имеет эволюционного смысла, если только индивид не путешествует со значительной долей близких родственников. Индивид, не путешествующий в таких обстоятельствах, должен предпочесть более безопасный вариант. Но в целом люди оценивают вероятность гибели не так, как актуарии; вместо этого они используют чувства, которые, вероятно, были сформированы естественным отбором. В результате их оценки риска могут быть неточными.
Подобно нам, Бодемер и соавторы придерживаются эволюционного подхода и утверждают, что "страх" рационален. Их анализ выходит за рамки нашего, поскольку рассматривает популяцию, структурированную по возрасту, но имеет некоторые серьезные ограничения. Одно из них — мера, которую они используют (человеко-годы), игнорирует структуру популяции и зависит от периода времени, за который она измеряется. Вместо того чтобы рассматривать рост всей популяции (например, всей территории США), эволюционный анализ должен рассматривать когорту, включающую тех индивидов, которые разделяют вторгающийся аллель. При этом мы используем геометрическую приспособленность, поскольку нас интересует совокупный риск, затрагивающий многих индивидов, несущих данный ген. Этот критический аспект риска отсутствует у Бодемера и соавторов, как и частота риска. В отличие от этого, мы вывели простое выражение, которое может быть использовано для сопоставления двух форм риска друг с другом, причем сопоставление зависит от доли родственников, погибших в результате события, связанного с совокупным риском.
Методы
Мы рассматриваем редкий мутантный аллель. Предполагаем, что индивиды, несущие этот аллель, имеют по одной его копии. Поведение такого индивида приводит к вероятности p того, что индивид погибнет в случае экологической катастрофы. Мы отслеживаем когорту таких индивидов во времени. Пусть N(t) — число членов когорты, присутствующих в момент времени t, что равно числу копий аллеля в момент времени t. Предполагается, что это число велико, так что мы можем усреднить демографическую стохастичность. Тогда между катастрофами изменение числа дается формулой:
$$ rac{dN}{dt} = RN(t) - M_D N(t) $$
Пусть x(t) = og (N(t)). Тогда между катастрофами:
$$ rac{dx}{dt} = R - M_D $$
Во время катастрофы происходит скачок вниз в N(t):
$$ N(t) ightarrow (1-p)N(t) $$
Таким образом:
$$ x(t) ightarrow x(t) + og (1-p) $$
(Заметим, что og (1-p) отрицательно.) Стохастический процесс x(t): t e 0 является стационарным во времени. Поскольку катастрофы происходят с интенсивностью ambda, согласно Уравнениям (7) и (9), этот процесс имеет среднее смещение:
$$ r = R - M_D + ambda og (1-p) $$
Долгосрочная скорость увеличения числа аллелей максимизируется максимизацией этого смещения. Поэтому мы принимаем r в качестве инвазивной приспособленности аллеля. (Заметим, что e^r — это геометрическая средняя скорость увеличения числа аллелей (например, [27]).) Используя соотношение M_C = ambda p, мы получаем:
$$ r = R M_D + (M_C/p) og (1-p) $$
$$ r = R (M_D + lpha M_C) $$
Доступность данных: Все данные, сгенерированные или проанализированные в ходе данного исследования, включены в данную опубликованную статью. Финансирование: SRXD получил поддержку от Senior Research Fellowship Королевского общества Leverhulme Trust. Информация об авторах: Авторы и их аффилиации: * Кафедра математики, Бристольский университет, Бристоль, Великобритания: Джон М. МакНамара * Центр экологии и охраны природы, Эксетерский университет, Пенрин, Великобритания: Саша Р. Х. Далл * Школа биологических наук, Бристольский университет, Бристоль, Великобритания: Аласдер И. Хьюстон Вклад авторов: * Дж. М. МакНамара: сформулировал модель и провел математический анализ. * С. Р. Х. Далл и А. И. Хьюстон: проанализировали литературу и несли основную ответственность за написание статьи. Контактное лицо: * John M. McNamara. Заявления об этике: Конфликты интересов: Конфликтов интересов нет. Дополнительная информация: Заявление издателя: Springer Nature сохраняет нейтралитет в отношении территориальных претензий, изложенных на картах и в институциональных аффилиациях. Права и разрешения: Открытый доступ: Эта статья распространяется по лицензии Creative Commons Attribution 4.0 International (CC BY 4.0), которая разрешает использование, обмен, адаптацию, распространение и воспроизведение на любом носителе или в любом формате при условии надлежащего указания авторства и источника, предоставления ссылки на лицензию Creative Commons и указания на внесение изменений. Изображения или другие материалы третьих сторон: Материалы третьих сторон, включенные в статью, подпадают под действие лицензии Creative Commons, если иное не указано в сноске к материалу. Если материал не включен в лицензию Creative Commons и предполагаемое использование не разрешено законодательством или выходит за рамки разрешенного использования, необходимо получить разрешение непосредственно у владельца авторских прав. Для просмотра копии лицензии посетите http://creativecommons.org/licenses/by/4.0/. Перепечатки и разрешения: О статье: McNamara, J.M., Dall, S.R.X. & Houston, A.I. Scales of risk and adaptive ‘dread’: an evolutionary theory of risk inflation. Sci Rep 15, 39211 (2025). https://doi.org/10.1038/s41598025190793 Скачать цитирование: Получено: 20 июня 2024 г. Принято: 05 сентября 2025 г. Опубликовано: 10 ноября 2025 г. Версия записи: 10 ноября 2025 г. DOI: https://doi.org/10.1038/s41598025190793 Поделиться статьей: Любой, кому вы отправите следующую ссылку, сможет прочитать этот контент: [Ссылка для общего доступа] Извините, ссылка для общего доступа в данный момент недоступна. Скопировать ссылку для общего доступа в буфер обмена: Предоставлено программой Springer Nature SharedIt для совместного использования контента.





