Интересное сегодня
Влияние западной диеты во время беременности на риск СДВГ и ...
Введение Огромное клиническое исследование, проведенное в Университете Копенгагена, проанализировало...
Аутизм и пищевые привычки: как связаны особенности поведения...
Аутизм и пищевые проблемы: глубокий анализ взаимосвязей Расстройства аутистического спектра (РА...
Исследование иллюзии визуального фантома и ее влияние на пос...
Введение Конструктивная природа зрительного восприятия движения ярко проявляется в явлениях восприни...
Влияние экономического положения и образа жизни на психическ...
Психическое здоровье и его значение Психическое здоровье является неотъемлемой частью общего благопо...
Синдром выгорания у преподавателей вузов: причины, последств...
Введение В последние годы растёт обеспокоенность состоянием здоровья преподавателей университетов. П...
Как финансовый стресс влияет на поведение детей: роль семейн...
Влияние финансовых трудностей на развитие детей Экономические трудности давно признаны фактором риск...

Влияние условий освещенности на зависимость частоты индуцированных транскраниальной электрической стимуляцией фосфенов
Введение
Многие аспекты нейронной обработки информации зависят от частотно-специфических колебаний в коре головного мозга. Это сделало привлекательной возможность исследования и/или манипулирования этими частотными нейронными функциями с помощью неинвазивных методов, таких как транскраниальная электрическая стимуляция (tES). Применение электрического тока к мозгу с помощью tES успешно модулирует когнитивные, сенсорные и моторные функции в зависимости от частоты по всей поверхности коры.
Хотя tES может модулировать кортикальную активность, она также может вызывать фосфены – зрительные ощущения, не связанные с внешними стимулами. Эти фосфены обычно считаются результатом электрической стимуляции сетчатки. Понимание биологических механизмов, ответственных за возникновение фосфенов, а также факторов окружающей среды, влияющих на их появление, имеет решающее значение, поскольку они могут искажать результаты исследований tES, интерпретации и вмешательства.
Например, порог возникновения фосфенов в настоящее время используется Международной комиссией по защите от неионизирующего излучения (ICNIRP) для установления ограничений воздействия. Эта информация может быть получена только при условии знания влияния частоты стимуляции и условий освещенности. Без этих знаний экспериментально полученные оценки порогов могут представлять собой лишь минимальные уровни стимуляции, необходимые для возникновения фосфенов в конкретных, нечувствительных сценариях, что ограничит способность защитных мер, основанных на этих данных, защищать от фосфенов в других ситуациях.
Последние исследования показывают, что наше понимание фосфенов может быть ограничено, особенно в отношении их связи с частотой стимуляции и условиями освещенности. Общепринято считать, что пороги возникновения фосфенов, индуцированных транскраниальной переменной стимуляцией (tACS), минимальны при частоте 20 Гц в условиях яркого освещения (фотопических) и при 10 Гц в полной темноте. Эта чувствительность близка к доминирующим частотам колебаний зрительной коры, наблюдаемым при соответствующих условиях освещенности. Эти выводы были представлены как свидетельство того, что tES, настроенная на доминирующую частоту кортикальных колебаний, может использоваться для максимальной модуляции кортикальной активности с аналогичными ритмами.
Однако недавние исследования предполагают, что в условиях сумеречного освещения (мезопических) фосфены tES индуцируются при значительно меньших токах при стимуляции 16 Гц, чем 20 Гц, что указывает на то, что 20 Гц не обеспечивает минимальный уровень стимуляции для возникновения фосфенов. Это вызывает сомнения в достоверности этих новых отчетов. Единственное доступное сравнительное исследование, явно тестирующее уровни порога фосфенов в мезопических условиях, – это исследование Шварца. Шварц сообщил о более низких порогах обнаружения фосфенов при стимуляции 20 Гц как при фотопических (8–9550 кд/м²), так и при мезопических (2.4 кд/м²) условиях. Однако эти выводы основывались только на одном испытуемом и использовали плохо контролируемые условия освещения. Это затрудняет выводы из такого сравнения.
В отличие от исследования Шварца, более поздние исследования использовали значительно большие выборки (24 или 22 участника в каждом из этих исследований) с точно контролируемым освещением (постоянное освещение 0.6 кд/м² по всему полю зрения). Более того, последовательность первоначального и репликационного исследования предполагает, что их результаты надежны. Этот вывод, однако, на первый взгляд, противоречит мнению о том, что максимальная чувствительность к tES достигается при частоте стимуляции, соответствующей доминирующей частоте кортикальных колебаний. То есть, в то время как доминирующие частоты кортикальных колебаний для фотопических и темных условий составляют примерно 20 и 10 Гц соответственно, а минимальный ток, необходимый для возникновения фосфенов, составляет 20 и 10 Гц соответственно, для мезопических условий соответствующей доминирующей частоты нет.
Одно из возможных объяснений более низких порогов при стимуляции 16 Гц заключается в том, что это представляет собой точку пересечения зависимостей порога и частоты стимуляции для фотопических и темных условий. То есть, поскольку условие слабого освещения представляет собой фотонную энергию, большую, чем в темноте, но меньшую, чем в фотопическом сценарии, оно может быть связано с порогом и частотой стимуляции как темных, так и фотопических условий. Для проверки этой гипотезы необходимо оценить зависимости порога и частоты стимуляции в каждом из условий: темном, мезопическом и фотопическом.
Различия в частотной зависимости, обнаруженные в индуцированных tES фосфенах в разных условиях освещенности, могут быть объяснены различиями в функциях временной чувствительности к контрасту – чувствительности зрительной системы к изменениям яркости во времени. Временная чувствительность к контрасту обычно измеряется с использованием однородного визуального стимула, который синусоидально изменяет яркость (от минимального до максимального значения) в зависимости от времени. Хотя этот стимул должен восприниматься как мерцание при больших уровнях контраста яркости, становится все труднее увидеть это мерцание по мере снижения контраста яркости. Однако пороговый уровень контраста яркости, при котором это мерцание едва заметно, также критически зависит от временной частоты стимула.
Исследования показали, что палочки и колбочки – фоторецепторы – имеют свои собственные функции временной чувствительности к контрасту, которые могут быть связаны с различиями в частотной зависимости, связанной с tES, обнаруженной в различных условиях освещенности. Палочки более чувствительны к стимуляции (и, следовательно, более вероятно, будут активированы) в более темных условиях, когда недостаточно стимула для активации колбочек. В темных и мезопических условиях временная чувствительность к контрасту в значительной степени обусловлена сигналами от палочек, особенно при воздействии стимулов, мерцающих с частотой 5–15 Гц, при этом наблюдается слабая или нулевая активация при 19–23 Гц. С другой стороны, колбочки более чувствительны к стимуляции в более ярких условиях, когда палочки насыщены и не вносят существенного вклада в визуальное восприятие. Временная чувствительность к контрасту в этих фотопических условиях имеет тенденцию к более высоким частотам, причем максимальная чувствительность наблюдается около 15–25 Гц, и нет возможности различать стимулы, мерцающие с частотой 80 Гц и выше. Если восприятие индуцированных tES фосфенов аналогично или функционально эквивалентно внешним мерцающим визуальным стимулам, то темные условия должны приводить к повышенной чувствительности к низкочастотной стимуляции, а яркие условия – к повышенной чувствительности к более высоким частотам.
Настоящее исследование было разработано для определения следующего. Во-первых, путем тестирования зависимости порога от частоты стимуляции в каждом из условий (темном, мезопическом и фотопическом), оно позволило выяснить, возникает ли общий порог обнаружения фосфенов при стимуляции 16 Гц, а не 20 Гц. Во-вторых, оно расширило результаты предыдущих исследований, которые обнаружили более низкие пороги при стимуляции 16 Гц в мезопических условиях (и которые увеличивались для более высоких и низких частот) по сравнению с темными и фотопическими условиями освещения. В-третьих, оно определило, можно ли объяснить зависимость порога фосфенов от частоты стимуляции в мезопических условиях пересечением зависимостей для темных и фотопических условий.
Методы
В исследовании приняли участие двадцать четыре здоровых добровольца (равное соотношение полов, возраст 20–40 лет, средний возраст 25.2 года, стандартное отклонение 5.4) после прохождения модифицированного чек-листа безопасности. Участники были исключены, если сообщали о каких-либо неврологических травмах или заболеваниях, наличии металлических имплантатов в голове или медицинских имплантатов в других частях тела, а также о некорректируемом нарушении зрения. Никто из участников не пользовался контактными линзами, а трое участников обычно носили очки, но снимали их во время тестирования, чтобы оправы не закрывали периферийное поле зрения. После ознакомления с процедурой исследования и потенциальными побочными эффектами tES, участники дали письменное информированное согласие до начала участия. Исследование проводилось в соответствии с Хельсинкской декларацией и было одобрено Комитетом по этике исследований человека Университета Вуллонгонга (одобрение № HE2017/454).
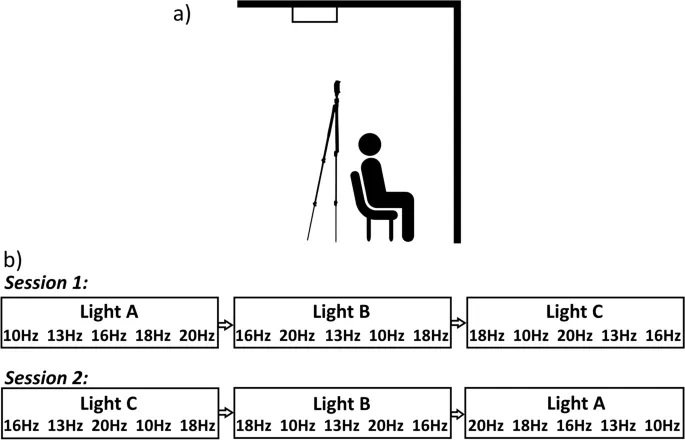
Пороги возникновения фосфенов определялись в зависимости от частоты стимуляции (10, 13, 16, 18 и 20 Гц) и условий освещенности (темнота, мезопическое, фотопическое) с использованием дизайна с повторными измерениями. Тестирование проводилось в два сеанса по 70 минут (в разные дни) примерно в одно и то же время суток и обычно в течение одной недели. Порядок сеансов, порядок монтажей tES в рамках этих сеансов, а также то, какой электрод был катодом или анодом в каждом монтаже, были сбалансированы между сеансами для всех участников. Выбор катода/анода чередовался между сеансами для каждого участника. Порядок условий освещенности рандомизировался для каждого участника с использованием системы латинского квадрата, как и порядок частоты стимуляции в каждом условии освещенности.
tES проводилась с использованием стимулятора Magstim NeuroConn DC Stimulator Plus MOP15EN01, который подавал синусоидальный постоянный ток без наращивания, то есть амплитуда стимуляции менялась по синусоиде от нуля до установленного тока, а затем обратно к нулю. Таким образом, полярность электродов не чередовалась. Ток подавался на кожу головы через электроды из проводящей резины (размеры: 3 × 4 см), помещенные на губки, пропитанные солевым раствором с гипоаллергенным амфотерным поверхностно-активным веществом, и закрепленные на FPz и Cz резиновыми ремнями. Такой монтаж электродов был выбран, поскольку он эффективно стимулирует сетчатку, одновременно гарантируя, что всё поле зрения участников не было перекрыто каким-либо оборудованием. Предыдущие исследования tES в условиях мезопического освещения последовательно выявляли наибольшую чувствительность при 16 Гц с использованием широкого диапазона монтажей, что предполагает, что выбор монтажа не оказывает существенного влияния на частотно-зависимую природу индуцированных tES фосфенов при условии адекватной стимуляции сетчатки.
Фотопическое освещение создавалось с помощью обычных потолочных люминесцентных ламп, расположенных вне прямого поля зрения участников. При таких условиях освещенности яркость для глаза измерялась как 77.1 ± 0.05 кд/м² с использованием люксметра J6523 Tektronix. Этот уровень освещенности был выбран из-за его применимости к повседневному опыту, поскольку он представляет собой яркость, обычно встречающуюся в офисных помещениях. Мезопическое освещение создавалось с помощью диммируемой светодиодной панели Neewer T120, освещающей область перед участником, что приводило к яркости для глаза, измеренной как 0.6 ± 0.05 кд/м². Этот уровень освещенности был выбран для соответствия предыдущим исследованиям и, таким образом, для обеспечения воспроизведения их результатов. В тестируемую комнату во время темного условия свет не проникал, что приводило к 0 кд/м². Чтобы предотвратить эффекты темновой адаптации во время проб в темном условии, освещение устанавливалось на 1.1 кд/м² при неактивном стимуляторе; освещение выключалось за 2 секунды до начала стимуляции и включалось сразу после ее окончания.
Участники сидели на стуле лицом к стене шириной 1.8 м и высотой 2.62 м, расположенной на расстоянии, обеспечивающем заполнение стены перед ними всем полем зрения, а освещение было настроено так, чтобы участник не видел теней. Когда участники располагались комфортно и электроды были установлены, им сообщали о фосфенах (их природе и том, что они могут воспринимать), пока кожа и волосы не были пропитаны солевым раствором из губок. Как только импеданс между электродами составлял 15 кОм или ниже (как указано стимулятором), освещение устанавливалось на 2 ± 0.05 кд/м², и участники знакомились с внешним видом фосфенов с помощью 10-секундной синусоидальной tES при 1000 мкА, сначала при 11 Гц, а затем при 22 Гц, чтобы продемонстрировать как визуальный вид фосфенов, так и то, как их внешний вид меняется простым изменением частоты стимуляции. Эти частоты были выбраны, чтобы избежать использования тех же частот стимуляции, что и в эксперименте.
После того, как участники ознакомились с фосфенами, определялись их пороги обнаружения фосфенов при каждой частоте стимуляции и условии освещенности. Им сообщали, когда начиналась и когда заканчивалась каждая стимуляция, но не сообщали частоту или интенсивность тока стимуляции. Стимуляции длились 5 секунд, и участникам было предложено держать глаза открытыми на протяжении всей стимуляции. На протяжении всего эксперимента участников спрашивали, насколько яркими кажутся фосфены по сравнению с фоновым освещением, и где фосфены появлялись в их поле зрения. Многие участники также спонтанно предоставляли информацию о своем опыте в интервале между пробами. Для выявления ложноположительных реакций при более низких уровнях интенсивности тока проводилось шесть контрольных стимуляций (по одной в каждом условии освещенности для каждого сеанса) с частотой, определенной заранее с помощью генератора случайных чисел на базе MATLAB. В этих контрольных пробах участнику давались все звуковые сигналы стимуляции (обычные нажатия кнопок на стимуляторе, а также словесные указания о начале и окончании стимуляции) без фактического генерирования электрического тока. Ни один из участников не сообщил о видении фосфенов во время каких-либо контрольных проб.
Пороги индукции фосфенов (в мкА) определялись для каждой частоты путем изменения интенсивности тока с использованием байесовской адаптивной процедуры лестничного поиска на основе QUEST в MATLAB PsychToolbox. tES, начинающаяся с 700 мкА, ограничивалась диапазоном от 25 до 1500 мкА. Шаг между возможными уровнями стимуляции составлял 25 мкА. Основываясь на системе быстрой оценки порога фосфенов, валидированной Mazzi et al., этот адаптивный метод измерения порога определял минимальную интенсивность тока, которая была значительно более вероятной, чем случайная, для вызова восприятия фосфенов. Каждый из двух сеансов предоставлял порог для каждого условия освещенности и частоты, а для каждой комбинации условия освещенности и частоты стимуляции средний порог по обоим сеансам принимался в качестве окончательного порога.
Статистический анализ
Поскольку значительные уровни асимметрии, эксцесса или гетерогенности дисперсии не были обнаружены, проводились параметрические анализы. Коррекция по Хюйн-Фельдту использовалась для учета нарушений сферичности (частота; взаимодействие частоты и условий освещенности), с указанием скорректированных степеней свободы.
Для оценки надежности порогов между двумя сеансами тестирования, для каждой комбинации условий освещенности (фотопическое, мезопическое, темное)/частоты (10, 13, 16, 18, 20 Гц) определялся коэффициент корреляции Пирсона (r). Для определения того, искажают ли результаты эффекты порядка, пороги были упорядочены хронологически, и использовался дисперсионный анализ с повторными измерениями, где порог был зависимой переменной, а порядок тестирования в пределах каждого условия освещенности (отдельно для каждого сеанса) и в рамках всего сеанса – независимыми переменными.
Для описания зависимостей между порогами фосфенов, условиями освещенности и частотой стимуляции использовался дисперсионный анализ с повторными измерениями, где порог был зависимой переменной, а условия освещенности и частота – независимыми переменными. При наличии значимых результатов данные далее анализировались с помощью t-критерия для повторных измерений с поправкой Бонферрони (условия освещенности: каждый уровень сравнивался с каждым другим уровнем; частота: каждый уровень сравнивался с каждым другим уровнем; взаимодействие: для каждой частоты каждый уровень условий освещенности сравнивался с каждым другим уровнем). Указаны скорректированные p-значения.
Для определения минимальных абсолютных порогов фосфенов по условиям освещенности использовался дисперсионный анализ с повторными измерениями, где условия освещенности были независимой переменной, а зависимой переменной – минимальный алгебраический порог по всем частотам для каждого условия освещенности отдельно. При наличии значимого главного эффекта проводились t-критерии с поправкой Бонферрони (условия освещенности: каждый уровень сравнивался с каждым другим уровнем). Указаны скорректированные p-значения.
Для определения того, можно ли адекватно объяснить зависимость порога фосфенов от частоты стимуляции в мезопических условиях суммированием зависимостей в темных и фотопических условиях, были рассчитаны регрессионные уравнения. Для получения оценки зависимости порога фосфенов от частоты стимуляции для каждого условия освещенности отдельно были проведены регрессионные анализы, где порог был зависимой переменной (нормализованный по всем протестированным частотам, в пределах каждого участника и условия освещенности отдельно), а частота – независимой переменной. Скорректированный информационный критерий Акаике (AICc) использовался для определения того, является ли линейная или квадратичная модель лучшей для каждого условия освещенности. Данные каждого участника использовались для всех частот и условий освещенности, что привело к 72 точкам данных на частоту.
Результаты
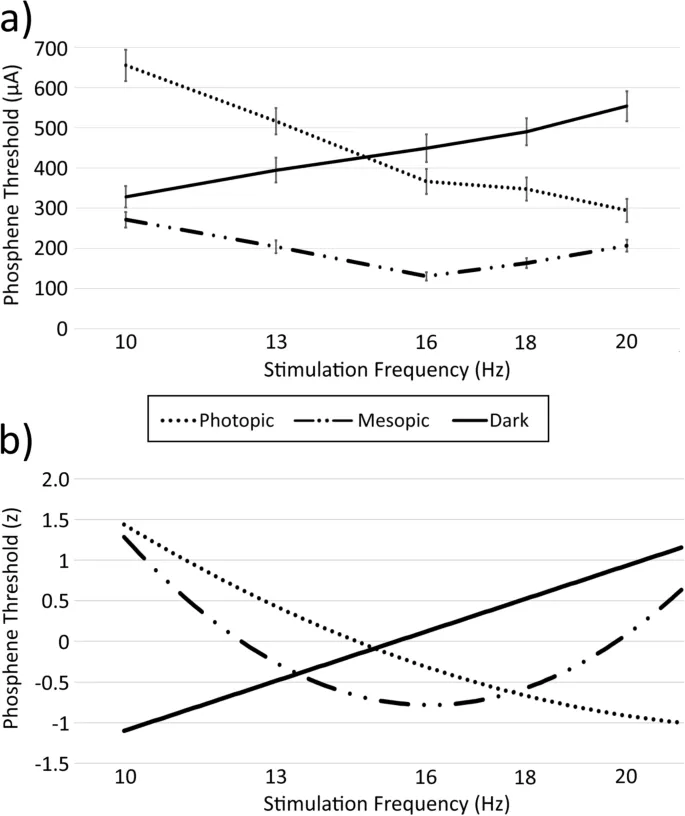
Пороги для каждой комбинации частоты/условий освещенности представлены на Рис. 2а и в Дополнительной таблице 1. Интерполированные регрессионные функции, связывающие порог с частотой, для каждого условия освещенности отдельно, представлены на Рис. 2b. Соответствующие средние значения и стандартные ошибки приведены в Таблице 1.
Пороги фосфенов были высоко надежны между двумя сеансами тестирования, с коэффициентами корреляции Пирсона (r) в диапазоне от 0.91 до 0.99 (все p < 0.999). На пороги фосфенов (для объединенных условий освещенности) также влияла частота (главный эффект: F(1.69,38.91) = 26.46, p < 0.001, η²p = 0.535). Частота с наименьшим порогом (16 Гц) была ниже, чем каждая другая частота (все p < 0.049), а частота с наибольшим порогом (10 Гц) была выше, чем каждая другая частота (p < 0.004). Среди остальных сравнений пороги для 13 Гц были выше, чем для 18 Гц (F(1, 23) = 10.34, p = 0.038, η²p = 0.310), но не отличались от 20 Гц (p = 0.795), а 18 Гц были ниже 20 Гц (F(1, 23) = 17.75, p = 0.003, η²p = 0.436).
Взаимодействие между частотой и условиями освещенности также было значимым (F(4.63,106.50) = 117.60, p < 0.001). Последующие анализы значимого взаимодействия показали следующее:
- 10 Гц стимуляция: Пороги были ниже как в мезопических (t(23) = 13.77, p < 0.001, Cohen’s d = 2.81), так и в темных (t(23) = 8.85, p < 0.001, d = 1.81) условиях, чем в фотопических, в то время как мезопические и темные условия не различались (t(23) = 2.25, p = 0.102, d = 0.46).
- 13 Гц стимуляция: Пороги были ниже в мезопических, чем в темных (t(23) = 7.58, p < 0.001, d = 1.55) и фотопических (t(23) = 12.31, p < 0.001, d = 2.51) условиях, и ниже в темных по сравнению с фотопическими условиями (t(23) = 4.73, p < 0.001, d = 0.97).
- 16 Гц стимуляция: Пороги были ниже в мезопических, чем в фотопических (t(23) = 9.64, p < 0.001, d = 1.97) и темных (t(23) = 10.27, p < 0.001, d = 2.10) условиях, в то время как темные и фотопические условия не различались (t(23) = 2.54, p = 0.055, d = 0.52).
- 18 Гц стимуляция: Пороги были ниже в мезопических, чем в фотопических (t(23) = 7.98, p < 0.001, d = 1.63) и темных (t(23) = 11.65, p < 0.001, d = 2.38) условиях, а фотопические были ниже темных (t(23) = 4.12, p = 0.001, d = 0.84).
- 20 Гц стимуляция: Пороги были ниже в мезопических, чем в фотопических (t(23) = 3.25, p = 0.009, d = 0.67) и темных (t(23) = 10.71, p < 0.001, d = 2.19) условиях, а фотопические были ниже темных (t(23) = 7.29, p < 0.001, d = 1.49).
Самые низкие пороги в каждом условии освещенности (по всем частотам) различались в зависимости от условий освещенности (главный эффект: F(2,46) = 34.84, p < 0.001, η²p = 0.602), с более низкими порогами, обнаруженными в мезопических условиях (при 16 Гц) по сравнению с темными (при 10 Гц; F(1, 23) = 54.88, p < 0.001, η²p = 0.705) и яркими (при 20 Гц; F(1, 23) = 66.42, p < 0.001, η²p = 0.743) условиями. Различий между минимальными порогами в ярком и темном условиях не обнаружено (p = 0.772).
Для темных условий линейная модель (AICc = 193.02) обеспечила лучший подбор, чем квадратичная модель (AICc = 194.34). Для мезопических условий квадратичная модель (AICc = 199.46) обеспечила лучший подбор, чем линейная модель (AICc = 289.71). Для фотопических условий квадратичная модель (AICc = 37.02) обеспечила лучший подбор, чем линейная модель (AICc = 64.34).
Регрессионные анализы (предсказывающие порог как функцию частоты) дали следующие уравнения (см. Рис. 2b):
- Темные условия:
Порог = −3.130 + 0.203 × Частота; adjR² = 0.650, p < 0.001. - Мезопические условия:
Порог = 13.682 − 1.800 × Частота + 0.056 × Частота²; adjR² = 0.634, p < 0.001. - Фотопические условия:
Порог = 6.587 − 0.655 × Частота + 0.014 × Частота²; adjR² = 0.905, p < 0.001.
Участники последовательно сообщали, что фосфены в мезопических и темных условиях увеличивались по яркости (по сравнению с фоном) с увеличением интенсивности стимуляции. В фотопических условиях участники последовательно сообщали, что мерцание фосфенов делало поле зрения темнее по сравнению с уровнями воспринимаемой яркости до стимуляции, где мерцание чередовалось между видимой вспышкой и предыдущим уровнем общей яркости. Это различие становилось более выраженным с увеличением интенсивности стимуляции. Четыре участника сообщили о видении цветных фосфенов, однако эти сообщения не были последовательными ни между этими участниками, ни внутри одного участника между сеансами.
Обсуждение
Целью исследования было определение зависимостей между порогами обнаружения фосфенов и как условиями освещенности, так и частотой стимуляции tES. Эти зависимости позволили нам проверить, подтверждается ли (1) недавние сообщения о повышенной чувствительности к индуцированным tES фосфенам при 16 Гц, в отличие от общепринятой точки зрения о максимальной чувствительности при 20 Гц, и (2) объясняется ли это кажущееся противоречие в литературе условиями освещенности.
Как видно из Рис. 2, каждое из условий освещенности имело уникальную зависимость порога фосфенов от частоты стимуляции, при которой пороги увеличивались с частотой в темном условии, снижались с частотой в фотопическом условии, и снижались с частотой от 10 до 16 Гц и увеличивались от 16 до 20 Гц в мезопическом условии. Соответственно, минимальные пороги в темных, мезопических и фотопических условиях были обнаружены при 10 Гц, 16 Гц и 20 Гц соответственно. Это демонстрирует, что как условие освещенности, так и частота стимуляции важны для определения минимального тока, необходимого для возникновения фосфенов. Корреляционный анализ показал последовательные пороги фосфенов между сеансами, указывая на то, что эффекты порядка и выбор катода или анода не оказали значительного влияния на результаты.
Самые низкие пороги в целом были обнаружены в мезопических условиях (при 16 Гц), при этом пороги были значительно выше как в темных (при 10 Гц), так и в фотопических (при 20 Гц) условиях. Это означает, что оценки порогов, полученные с использованием стандартных фотопических или темных условий, независимо от частоты, будут завышать ток, необходимый для возникновения фосфенов. Следовательно, руководства, использующие обнаружение фосфенов для установления ограничений воздействия на основе данных, полученных в темных или фотопических условиях (например, ICNIRP), могут недооценивать влияние электрического тока на нейронные процессы (на 56–60%). Важно отметить, что, хотя такие руководства обычно полагаются на исследования с использованием магнитных полей (а не tES) для индукции фосфенов, в обоих случаях причиной фосфена является ток, протекающий через нервную ткань, который стимулирует те же физиологические процессы. Следовательно, настоящие результаты применимы также к исследованиям, использующим магнитные поля для индукции фосфенов, и, таким образом, к руководствам по воздействию низкочастотных электромагнитных полей.
Настоящие данные также разрешают кажущееся противоречие между недавними исследованиями, которые обнаружили пороги обнаружения фосфенов при 16 Гц, и исследованиями, сообщающими пороги при 10 или 20 Гц. То есть, настоящие результаты демонстрируют, что мезопические условия обеспечивают максимальную чувствительность при 16 Гц, в то время как темные и фотопические условия (аналогичные тем, что использовались в прошлых исследованиях) обеспечивают максимальную чувствительность при tES 10 Гц и 20 Гц соответственно. Таким образом, никакого противоречия нет, только предсказуемые различия из-за использования различных условий освещенности. В целом, результаты данного исследования, касающиеся зависимости от освещенности и частоты, согласуются с данными по различным формам tES, будь то tACS или синусоидальная tDCS. Хотя существуют различия в уровнях тока, необходимых для индукции фосфенов в разных исследованиях, это, вероятно, является результатом различных методологических выборов, поскольку существует множество переменных, которые могут изменять этот порог. Например, даже при использовании одного и того же монтажа на одной и той же выборке, значения порогов могут варьироваться в зависимости от размера, формы и площади поверхности электродов, материала, из которого изготовлен электрод, и выбранной среды проводимости (например, проводящий гель, губки, пропитанные электролитом). Изменение любой из этих переменных изменит характеристики объемного проведения всего контура, что приведет к различным уровням плотности тока в сетчатке. В результате, хотя сравнение порогов между исследованиями имеет небольшое значение, результаты, касающиеся частоты и освещенности, остаются последовательными, несмотря на любые вариации в методологии стимуляции.
Хотя соблазнительно предположить, что во время tES в мезопических условиях задействованы иные физиологические процессы по сравнению с темными и фотопическими условиями, более простое объяснение может быть достаточным для объяснения результатов. Как видно из Рис. 2, форма оценочного распределения порогов в мезопических условиях соответствовала темным условиям на частотах ниже примерной точки пересечения 16 Гц, а также соответствовала фотопическим условиям на частотах выше точки пересечения 16 Гц. В совокупности это позволяет предположить, что мезопические условия могут просто представлять собой комбинацию физиологических процессов, обычно задействованных в каждом из темных и фотопических условий.
В соответствии с этой гипотезой, существуют данные, свидетельствующие о том, что наблюдаемая частотная зависимость в исследованиях индуцированных tES фосфенов может быть объяснена различиями в относительной активности видения, основанного на палочках и колбочках. Клетки, связанные со зрением палочек, которые готовы реагировать в темных условиях, наиболее чувствительны к стимуляции около 10 Гц, в то время как клетки, связанные со зрением колбочек, которые готовы реагировать в фотопических условиях, наиболее чувствительны к стимуляции около 20 Гц. Само по себе это не объясняет величину снижения порога в мезопических условиях (60% и 56% по сравнению с темными и фотопическими условиями соответственно), особенно учитывая, что 16 Гц далеко не идеальная частота стимуляции как для клеток, связанных с палочками, так и для колбочек. Однако, в сочетании с известными данными о задержке обработки сигналов палочек и колбочек, это представляется жизнеспособной гипотезой. То есть, существует задержка между обработкой сигналов палочек и колбочек в мезопических условиях, но по мере достижения частоты стимуляции примерно 15 Гц палочки и колбочки начинают работать в фазе, что увеличивает сигнал как в палочках, так и в колбочках и усиливает воспринимаемость стимуляции. Эта критическая частота 15 Гц также приближается к точке пересечения регрессионных оценок для темных и фотопических условий (см. Рис. 2b), что указывает на то, что механизм фазовой задержки палочек и колбочек может быть причиной более низких общих порогов на близкой частоте 16 Гц в мезопических условиях. Дальнейшие исследования потребуются для проверки этой гипотезы.
Хотя высокие уровни тока могут вызывать дискомфорт или боль в месте стимуляции, эти эффекты обычно обнаруживаются при уровнях стимуляции, превышающих максимальные, использованные в данном исследовании. Максимальный уровень стимуляции (1500 мкА) в настоящем исследовании был выбран, чтобы избежать таких побочных эффектов. Один участник сообщил о неприятном ощущении, похожем на зуд, при уровнях стимуляции выше 900 мкА, однако ощущение немедленно прекращалось после прекращения стимуляции. Несмотря на многократные расспросы во время каждого сеанса тестирования, ни один другой участник не сообщил о каких-либо негативных побочных эффектах, ни во время, ни после стимуляции. Действительно, поскольку в исследовании намеренно использовались низкие уровни тока для определения порогов, это уменьшило возможность получения значимой информации об опыте восприятия фосфенов в более общем плане, что могло бы помочь пролить свет на лежащие в основе физиологические механизмы, ответственные за индукцию фосфенов. Особый интерес представляет степень, в которой фосфены воспринимались в хроматическом (в отличие от ахроматического) цвете, поскольку это могло бы свидетельствовать о релятивных механистических ролях палочек и колбочек в зависимости от частоты и условий освещенности. Однако, учитывая низкие уровни тока, использованные в исследовании, только четыре участника сообщили о видении хроматического цвета, и сообщения не были последовательными ни между этими участниками, ни внутри одного участника между сеансами. Поэтому мы не считаем, что эти анекдотические сообщения достаточны для интерпретации.
Заключение
Настоящее исследование показало, что кажущееся противоречие в литературе относительно частоты стимуляции tES и порога обнаружения фосфенов было обусловлено различными условиями освещенности, использованными в прошлых исследованиях. То есть, в то время как пороги в темных и фотопических условиях минимальны при стимуляции 10 Гц и 20 Гц соответственно, они не отражают общие пороги, которые возникают при 16 Гц в мезопических условиях. Величина завышения порога была очень велика (60% и 56% для темных и фотопических условий соответственно) и, следовательно, важна для применения исследований tES. Физиологические соображения предполагают, что более низкие пороги в мезопических условиях, и особенно при стимуляции 16 Гц, могут быть связаны с участием как палочек, так и колбочек фоторецепторов, но для подтверждения этого требуются дальнейшие исследования. Важно отметить, что наше исследование также показывает (впервые), что темные, мезопические и фотопические условия освещенности имеют свои уникальные зависимости порога фосфенов от частоты стимуляции.
Данные
Набор данных, полученный в результате этого эксперимента, доступен у соответствующего автора по обоснованному запросу.
Ссылки
Скачать ссылки.
Финансирование
Спонсор гранта: Центр исследований здоровья населения в области электромагнитной энергии, грант № APP1060205.
Информация об авторах
- Авторы и их места работы:
- Школа психологии, Университет Вуллонгонга, Вуллонгонг, Австралия
- Иэн Эванс, Стивен Палмисано и Родни Дж. Крофт
- Институт исследований здравоохранения и медицины Иллаварра, Университет Вуллонгонга, Вуллонгонг, Новый Южный Уэльс, 2522, Австралия
- Иэн Эванс, Стивен Палмисано и Родни Дж. Крофт
- Австралийский центр исследований биоэффектов электромагнитных полей, Вуллонгонг, Австралия
- Иэн Эванс и Родни Дж. Крофт
- Центр исследований здоровья населения в области электромагнитной энергии, Университет Монаша, Мельбурн, Австралия
- Иэн Эванс и Родни Дж. Крофт
Вклад авторов
Все авторы спланировали исследование. И. Э. собрал и обработал данные. Все авторы участвовали в интерпретации результатов, написании рукописи и одобрили окончательную версию.
Корреспондирующий автор
Переписка с: Иэн Эванс.
Заявления об этике
- Конкурирующие интересы: Авторы заявляют об отсутствии конкурирующих интересов.
Дополнительная информация
Примечание издателя: Springer Nature остается нейтральной в отношении юрисдикционных претензий, касающихся опубликованных карт и институциональных принадлежностей.
Дополнительные материалы
Дополнительная таблица 1.
Права и разрешения
Открытый доступ: Эта статья распространяется под лицензией Creative Commons Attribution 4.0 International License, которая разрешает использование, обмен, адаптацию, распространение и воспроизведение на любом носителе или в любом формате, при условии, что вы укажете соответствующий авторский кредит и источник, предоставите ссылку на лицензию Creative Commons и укажете, были ли внесены изменения. Изображения или другие материалы третьих лиц, включенные в статью, включены в лицензию Creative Commons статьи, если иное не указано в кредитной строке к материалу. Если материал не включен в лицензию Creative Commons статьи, и ваше предполагаемое использование не разрешено законом или превышает разрешенное использование, вам потребуется получить разрешение напрямую от владельца авторских прав. Чтобы просмотреть копию этой лицензии, посетите http://creativecommons.org/licenses/by/4.0/.
Перепечатки и разрешения
Об этой статье
Цитировать эту статью:
Evans, I., Palmisano, S. & Croft, R.J. Effect of ambient lighting on frequency dependence in transcranial electrical stimulation-induced phosphenes. Sci Rep 12, 7775 (2022). https://doi.org/10.1038/s41598-022-11755-y
Скачать цитирование.
Получено: 10 декабря 2021 г.
Принято: 21 апреля 2022 г.
Опубликовано: 11 мая 2022 г.
Версия для печати: 11 мая 2022 г.
DOI: https://doi.org/10.1038/s41598-022-11755-y
Поделиться этой статьей:
Любой, с кем вы поделитесь следующей ссылкой, сможет прочитать этот контент:
Получить ссылку для обмена.
Извините, ссылка для обмена в настоящее время недоступна для этой статьи.
Скопировать ссылку для обмена в буфер обмена.
Предоставлено инициативой Springer Nature SharedIt по обмену контентом.
На эту статью ссылаются
- Effects of different transcranial magnetic stimulation coil types on phosphene thresholds and their association with motor cortex excitability
- Halit Fidancı, Halil Can Alaydın, Zülfikar Arlıer
- BMC Neuroscience (2025)
- Exploring the correlation and causation between alpha oscillations and onesecond time perception through EEG and tACS
- Ehsan Mokhtarinejad, Mahgol Tavakoli, Amir Hossein Ghaderi
- Scientific Reports (2024)