Интересное сегодня
Использование ограничительных мер при кормлении через назога...
Введение Применение ограничительных мер для установки назогастрального зонда (НГЗ) с целью кормления...
Влияние угрозы на сложное принятие решений: данные из виртуа...
Влияние угрозы на сложное принятие решений Некоторые из самых сложных решений в жизни принимаются в ...
Как ваше лицо может стать новым жизненным показателем?
Введение Недавно исследование, опубликованное в журнале The Lancet Digital Health, представил...
Age-Related Predictive Processes in Language Comprehension
Введение в предсказательные процессы в языке Предположение о том, что люди, понимающие язык, использ...
Как физическая активность в свободное время и осознанность с...
Введение Академическое выгорание — это состояние эмоционального истощения, вызванное учебными задача...
Калибровочный эффект в визуомоторных задачах: исследование н...
Введение Люди часто демонстрируют менее точные результаты при первом выполнении задачи в серии попыт...




Влияние рецепторов окситоцина на развитие и поддержание социального поведения у данио-рерио (Danio rerio)
Рыбы данио-рерио (Danio rerio) — высокосоциальные лучепёрые рыбы, являющиеся превосходной моделью для изучения социального поведения. Нейропептид окситоцин ассоциируется с различными социальными поведением, а также с расстройствами, приводящими к нарушениям социального взаимодействия, такими как расстройства аутистического спектра. Однако неизвестно, как сигнализация окситоциновых рецепторов влияет на развитие и кинетику проявления социального поведения.
Введение
Многие виды, включая человека, живут в группах для повышения своей приспособленности — общей репродуктивной успешности за всю жизнь. Жизнь в социальном контексте предлагает множество преимуществ, таких как улучшенное обнаружение хищников и пищи1, доступность половых партнеров, снижение потребления энергии2, а также возможность учиться жизненно важным поведенческим реакциям у сородичей3. Для оптимизации совместного проживания в группе развились различные формы социального поведения.
Данио-рерио (Danio rerio), мелкая лучепёрая рыба, является мощной животной моделью, используемой в биомедицинских исследованиях, включая разработку лекарств4, биологию развития5,6 и нейробиологию7,8. Кроме того, данио-рерио демонстрируют разнообразные поведенческие реакции, включая избегание9, поиск пищи и охоту10, реакции на стресс11 и различные формы социальности12,13,14,15,16. Примеры социальности включают брачное поведение, агрессивное поведение и другие более простые поведенческие реакции, возникающие в группах. Данио-рерио предпочитают плавать в сплоченных стаях, тенденция, которая развивается в течение первых недель жизни17. Плавание в непосредственной близости к сородичам, также называемое социальной предпочтительностью, начинает развиваться уже с 1–2 недель после оплодотворения (wpf)18,19. Хотя развитие стайного поведения коррелирует с возрастными изменениями в дофаминергической и серотонинергической системах20, механизмы, лежащие в основе развития и поддержания социальной предпочтительности и стайного поведения, недостаточно изучены.
Нонапептид окситоцин — это высококонсервативный нейропептид, присутствующий у человека и с незначительными изменениями у большинства других животных21. Ортологичный ген окситоцина у данио-рерио (обозначается как Oxt) отличается от человеческого окситоцина всего двумя аминокислотами21. Помимо роли в родах22 и лактации23, окситоцин упоминается в контексте консолидации памяти24 и болевого поведения25,26,27,28. Уровни окситоцина в мозге также влияют на тревожность у мышей29,30,31, крыс32,33 и людей34,35. Кроме того, связь между окситоцином и социальным поведением была продемонстрирована в многочисленных исследованиях36,37,38, и рецептор окситоцина человека может влиять на социальные черты, затронутые при нейроонтологических расстройствах, таких как расстройства аутистического спектра39,40.
Социальная изоляция в течение нескольких недель влияет на различные поведенческие реакции грызунов, включая тревожность41, агрессию42 и депрессивное поведение43. Gilles и Polston44 сообщили, что двухнедельная изоляция крыс, начавшаяся после отъема от матери (постнатальный день 21), привела к усилению про- и антисоциального поведения. Они также описали обратную корреляцию между количеством окситоцинергических нейронов в передней парвицеллярной области гипоталамического паравентрикулярного ядра (PVN) и уровнем антисоциального поведения44. После длительной социальной изоляции применение окситоцина показало снижение депрессивного поведения у луговых полевок43 или агрессивного поведения у самцов мышей42. Более того, различные исследования с использованием крыс показали, что количество окситоцинергических нейронов в PVN уменьшается45 (постнатальный день 38–48), увеличивается46 (постнатальный день 74) или остается неизменным44 (постнатальный день 103–107) после социальной изоляции. Сообщалось также, что изоляция снижает активность рецепторов окситоцина (OTR) в прилежащем ядре крыс46 и влияет на экспрессию OTR в определенных областях мозга в разной степени: четырехнедельная социальная изоляция луговых полевок привела к снижению гипоталамической экспрессии OTR47, тогда как шести- и восьминедельная социальная изоляция не изменила экспрессию OTR в гиппокампе мышей48.
У мышей рецептор окситоцина экспрессируется в различных областях мозга, включая гиппокамп, миндалевидное тело, супрахиазматическое ядро и прелимбическую кору49. Поскольку лучепёрые рыбы, такие как данио-рерио, пережили раунд полного удвоения генома примерно 320–350 миллионов лет назад50, они обладают двумя ортологичными рецепторами окситоцина: рецептором окситоцина (Oxtr) и рецептором окситоцина подобным (Oxtrl). Несколько исследований51,52,53 изучали, как рецепторы окситоцина влияют на социальное распознавание или социальную предпочтительность у взрослых данио-рерио путем блокирования обоих рецепторов (Oxtr и Oxtrl) антагонистами51,52 или нокаута гена oxtr53. Однако часто эти исследования приходили к противоречивым выводам, подчеркивая сложность эффектов окситоцина. Дискретное влияние Oxtr или Oxtrl на развитие и поддержание социального поведения у данио-рерио, выращенных в социальных условиях или в изоляции, остается невыясненным.
Основной целью данного исследования было использование сгенерированных с помощью CRISPR/Cas9 нокаут-линий oxtr и oxtrl для изучения роли рецепторов окситоцина в развитии и поддержании социального поведения, в частности, социальной предпочтительности и стайного поведения у данио-рерио (Danio rerio).
Результаты
Для исследования роли рецепторов окситоцина в развитии и поддержании социального поведения мы сравнили рыб с нокаутом oxtr−/− и oxtrl−/− и их дикие типы (wildtype) по двум поведенческим реакциям: социальная предпочтительность (оценивает предпочтение рыбы социальной зоны по сравнению с не социальной) и стайное поведение.
Социальная предпочтительность
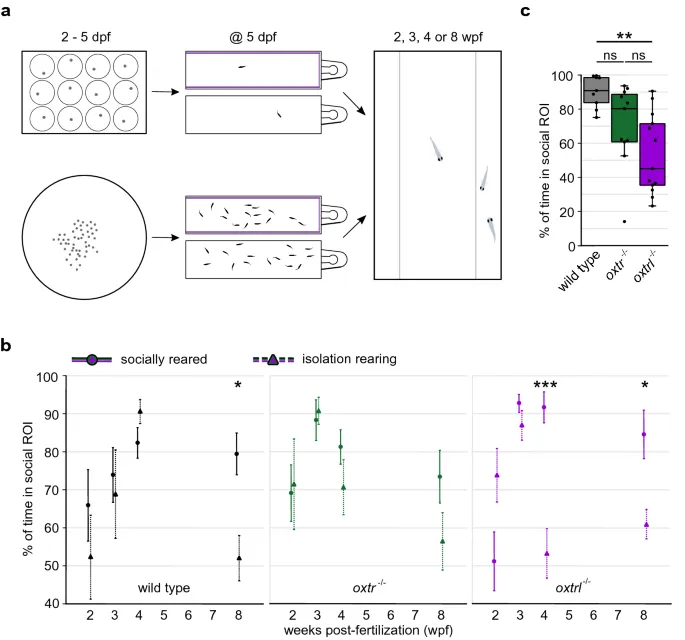
Для измерения социальной предпочтительности индивидуальных личинок данио-рерио на разных стадиях развития помещали в прямоугольный резервуар с одной зоной, где сородичей можно было наблюдать через прозрачную стеклянную стенку (Рис. 1а). После периода адаптации в стимулирующую зону резервуара добавляли двух сородичей, и наблюдалось поведение экспериментальной рыбы. Мы измеряли время, проведенное вблизи сородичей (социальная область интереса), как показатель социальной предпочтительности. У диких данио-рерио эта предпочтительность развивалась постепенно в течение первых 4 wpf и сохранялась до 8 wpf (Рис. 1b,c). Увеличение социальной предпочтительности, наблюдаемое между 2 и 4 wpf, было сопоставимо с результатами предыдущих исследований19,54, хотя и не было статистически значимым (p = 8.03 × 10−1). Напротив, у рыб с нокаутом oxtr−/− наблюдался ранний пик максимального социального поведения в 3 wpf. В отличие от диких рыб, эта высокая социальность не поддерживалась, а значительно (p = 4.95 × 10−2) снижалась в период с 4 по 8 wpf (Рис. 1b,c). Развитие социальной предпочтительности также было существенно (p = 2.09 × 10−4) изменено у рыб с нокаутом oxtrl−/−, которые также демонстрировали раннее начало социальной предпочтительности, преждевременно достигая пика в 3 wpf (Рис. 1b,c). Кроме того, максимальный уровень социальной предпочтительности, демонстрируемый обеими линиями нокаут-рыб по рецепторам окситоцина, был существенно выше, чем максимальный уровень, демонстрируемый дикими рыбами.
Социальная изоляция, как сообщается, влияет на социальное поведение у разных видов55,56,57,58. Поэтому мы далее изучили влияние содержания в изоляции на развитие и проявление социальной предпочтительности как у диких, так и у рыб с нокаутом oxtr−/− и oxtrl−/−. Мы индивидуально содержали (в отсутствие сородичей) диких, oxtr−/− и oxtrl−/− рыб с 2 дней после оплодотворения (dpf) до дня эксперимента (Рис. 2а). У диких рыб содержание в изоляции не оказало значительного влияния (p = 3.39 × 10−1) на кинетику начала социальной предпочтительности, но резко снизило поддержание социальной предпочтительности, обычно наблюдаемое в 8 wpf (Рис. 2b). Действительно, в 8 wpf, рыбы, выращенные в изоляции, не проявляли значимой (p = 3.26 × 10−1) социальной предпочтительности. В отличие от диких рыб, выращенных в социальных условиях, увеличение социальной предпочтительности с 2 до 4 wpf было значительным после содержания в изоляции (p = 1.22 × 10−2). Однако прямое сравнение диких рыб, выращенных в социальных условиях и в изоляции, одного возраста не выявило статистически значимых различий в уровнях социальной предпочтительности в 2–4 wpf. Содержание в изоляции оказало аналогичное воздействие на рыб oxtr−/−: изоляция не изменила кинетику начала социальной предпочтительности, но привела к ускоренному снижению социальной предпочтительности, измеренной в 8 wpf (Рис. 2b). Интересно, что рыбы oxtrl−/− демонстрировали ту же закономерность, но ускоренное снижение социальной предпочтительности после содержания в изоляции было преждевременно заметно в 4 wpf (Рис. 2b и c). Кроме того, ОТ-ПЦР экспрессии гена oxt у рыб, выращенных в социальных условиях и в изоляции, выявила, что изоляция привела к статистически значимому снижению уровней oxt в 8 wpf, но не в 4 wpf (см. Дополнительный Рис. S1c). Экспрессия генов oxtr и oxtrl, однако, не была затронута содержанием в изоляции (Дополнительный Рис. S1d и S1e). Эти данные показывают, что социальная предпочтительность развивается после содержания в изоляции у диких, oxtr−/− и oxtrl−/− рыб, но эта социальная предпочтительность не поддерживается. Снижение социальной предпочтительности, вызванное содержанием в изоляции, проявляется раньше у рыб oxtr−/− и oxtrl−/− (Рис. 2c).
Анализ скорости плавания выявил значительные различия между дикими и мутантными рыбами в 2 и 3 wpf (см. Дополнительный Рис. S3a и S3b); эти различия не коррелировали с описанными изменениями в развитии социальной предпочтительности. Например, как 2-, так и 3-недельные рыбы oxtrl−/− плавали значительно быстрее, чем дикие рыбы, но демонстрировали значительно усиленную (3 wpf) или сниженную (2 wpf) социальную предпочтительность. Другой пример — значительно сниженная социальная предпочтительность у 4-недельных рыб oxtrl−/− или oxtr−/−, выращенных в изоляции, со средней скоростью плавания, сравнимой с дикими данио-рерио. В совокупности эти данные указывают на то, что изменения в скорости плавания, вызванные потерей одного из рецепторов окситоцина, не дают прямого объяснения наблюдаемым изменениям в социальном поведении.
Стайное поведение
Мы также исследовали другую форму социального поведения: стайное поведение. Для измерения стайного поведения мы использовали установку, позволяющую 20 данио-рерио свободно перемещаться в круглом резервуаре (см. Дополнительный Рис. S2a) в течение 30 минут. Мы тестировали выращенных в социальных условиях диких, oxtr−/− и oxtrl−/− рыб в возрасте 4 и 8 wpf (более молодые рыбы не могли быть надежно отслежены из-за их малого размера). Для количественной оценки стайного поведения мы анализировали расстояние до ближайшего соседа, межвидовое расстояние и расстояние до самого дальнего соседа каждой рыбы, а также суммарное расстояние стаи и параметр поляризации, полученный из анализа главных компонент индивидуальных видео (см. «Материалы и методы»). В 4 wpf ни один из нокаутов рецепторов окситоцина не демонстрировал характеристик стайного поведения, которые существенно отличались бы от соответствующих диких контролей (p(oxtr+/+ vs. oxtr−/−) = 4.74 × 10−1 (a), 4.09 × 10−1 (b), 4.74 × 10−1 (c) и 8.88 × 10−1 (d), p(oxtrl+/+ vs. oxtrl−/−) = 3.17 × 10−1 (a), 2.59 × 10−1 (b), 1.29 × 10−1 (c) и 4.80 × 10−1 (d)) (Рис. 3a–d). Однако в 8 wpf обе нокаут-линии демонстрировали существенные различия в параметрах стайного поведения, все из которых соответствовали общему фенотипу менее сплоченной стаи (p(oxtr+/+ vs. oxtr−/−) = 1.64 × 10−3 (a), 1.38 × 10−2 (b), 1.91 × 10−2 (c) и 4.95 × 10−2 (d), p(oxtrl+/+ vs. oxtrl−/−) = 4.31 × 10−2 (a), 2.01 × 10−2 (b), 1.91 × 10−2 (c) и 4.95 × 10−2 (d)) (Рис. 3a–d). Суммарное расстояние стаи не изменилось ни у oxtr−/−, ни у oxtrl−/− (см. Дополнительный Рис. S2b). Кроме того, нокаут одного из рецепторов окситоцина не привел к увеличению тревожности, проявляющейся в смелости (см. Дополнительный Рис. S2c) и замирании (см. Дополнительный Рис. S3c и S3d). В совокупности эти два эксперимента показали, что оба рецептора окситоцина играют ключевую роль в развитии социальной предпочтительности и сплоченности стаи.
Обсуждение
Многие исследования59,60,61 продемонстрировали связь между окситоцином и социальным поведением, но точная роль двух рецепторов окситоцина в развитии и поддержании социального поведения оставалась неясной. Наши данные указывают на то, что предпочтение к сородичам постепенно развивается у диких данио-рерио в течение первых нескольких недель, достигая максимальной социальной предпочтительности в 4 wpf, которая затем поддерживается на высоком уровне по крайней мере до 8 wpf. Если один из рецепторов окситоцина (Oxtr или Oxtrl) был удален, развитие социальной предпочтительности ускорялось, преждевременно достигая максимального уровня в 3 wpf. Хотя расстройства аутистического спектра вызваны множеством факторов, таких как мутации в гене Fragile × mental retardation 1 (FMR1)62, интраназальное введение окситоцина может облегчить некоторые социальные нарушения у аутичных пациентов40, и, кроме того, специфические однонуклеотидные полиморфизмы (SNP) гена OXTR человека39 часто встречаются у популяции с РАС и описаны как потенциальный генетический фактор риска для РАС63. Мы показали здесь, что развитие социальной предпочтительности затрагивается рецепторами окситоцина у отдаленно родственного позвоночного — данио-рерио. Интересно отметить, что недавнее исследование показало, что развитие социальной предпочтительности также ускоряется у данио-рерио fmr1−/−64.
Содержание данио-рерио в изоляции не повлияло на временную динамику развития социальной предпочтительности, но существенно повлияло на ее поддержание. Недавнее исследование Tunbak и коллег65 описало снижение социальной предпочтительности примерно у 40% рыб после содержания в изоляции. Они использовали диких рыб генетического фона AB и поведенческую камеру, описанную ранее19, которая, в отличие от используемой здесь камеры, не обеспечивает одновременного визуального доступа к обеим зонам стимуляции. К сожалению, возраст рыб не был точно определен, но они классифицировали экспериментальных рыб как «ювенильных», предполагая возраст от 30 до 89 dpf66. В наших экспериментах рыбы, выращенные в изоляции, демонстрировали социальную предпочтительность, сравнимую с рыбами, выращенными в социальных условиях, в возрасте от 14 до 28 dpf, но они были менее социальны после 8 недель изоляции. Эти данные предполагают, что сниженная социальная предпочтительность является реакцией на периоды изоляции, превышающие 28 dpf, но различия в дизайне эксперимента (различные среды содержания в изоляции, поведенческие камеры, условия освещения, количество стимулирующих рыб) могут влиять на результаты тестов социальной предпочтительности.
Индуцированное изоляцией снижение социальной предпочтительности, которое мы наблюдали у диких данио-рерио в нашем исследовании, было ускорено у обоих рецепторов окситоцина нокаутов. ОТ-ПЦР экспрессии гена oxt у рыб, выращенных в социальных условиях и в изоляции, выявила, что изоляция привела к статистически значимому снижению уровней oxt в 8 wpf, но не в 4 wpf (см. Дополнительный Рис. S1c). Кроме того, в отличие от рыб, выращенных в социальных условиях, рыбы, выращенные в изоляции, не проявляли статистически более высокой экспрессии oxt с увеличением возраста. Эти данные согласуются с наблюдаемыми поведенческими изменениями после содержания в изоляции. Экспрессия генов oxtr и oxtrl, однако, не была затронута содержанием в изоляции (см. Дополнительный Рис. S1d и S1e), что противоречит исследованию, сообщающему о сниженной экспрессии рецептора окситоцина (OTR) у луговых полевок после четырех недель социальной изоляции47. Помимо сниженной экспрессии oxt у изолированных рыб, другое возможное объяснение драматически сниженного уровня социальной предпочтительности в 8 wpf может быть связано с регуляцией вниз гена pth2, поскольку изоляция также приводит к снижению экспрессии этого нейропептида67.
Наши результаты отличаются от работы Landin и коллег, которые тестировали социальную предпочтительность у взрослых и 3-недельных данио-рерио после введения антагониста рецептора окситоцина L368,899, специфического ингибитора как Oxtr, так и Oxtrl52. На обеих стадиях развития они обнаружили снижение социальной предпочтительности. В отличие от исследования Landin и др., предыдущее исследование Zimmermann и коллег не обнаружило значительного снижения социальной предпочтительности после интраперитонеального введения L368,899. Противоречивые результаты двух исследований были приписаны использованию разных концентраций антагониста — 0,01 нг/г51 и 100 мкг/г52. В отличие от этих двух исследований, мы использовали генетические мутанты, сгенерированные с помощью CRISPR/Cas9, с отсутствием только одного из двух рецепторов окситоцина, что позволило нам проанализировать их различные индивидуальные роли на протяжении развития. Оба рецептора влияли на развитие социальной предпочтительности, но, в отличие от oxtrl−/−, рыбы oxtr−/− не поддерживали высокий уровень социальной предпочтительности с возрастом. Wircer и коллеги68 описали субпопуляцию окситоцинергических нейронов, расположенных в заднем бугорке и экспрессирующих oxtr. Абляция этих нейронов привела к снижению социальной предпочтительности у взрослых данио-рерио68. Интересно, что исследование Ribeiro и коллег53 показало нарушение социального распознавания, но неизменную социальную предпочтительность у линии данио-рерио с нокаутом одного рецептора окситоцина (oxtrwz16/wz16). Различия в результатах между Ribeiro и др. и нашим исследованием могут быть связаны с различиями в экспериментальных условиях или возрасте, поскольку они использовали другую поведенческую камеру для теста социальной предпочтительности и рыб в возрасте от трех до шести месяцев. В их исследовании не тестировались рыбы с нокаутом oxtrl. В другом исследовании Ribeiro и коллеги описали незначительную тенденцию к снижению социальной предпочтительности у взрослых диких и нокаутных рыб oxtr после содержания в присутствии сородичей с нокаутом oxtr69. В наших экспериментах мы содержали диких рыб с дикими, а нокаутных рыб — с нокаутами. Различные исследования70,71,72,73 с мышами с нокаутом Oxtr показали снижение социального поведения и нарушение социального распознавания с усилением агрессии. Примечательно, что дефицит социального поведения также наблюдался у гетерозиготных мышей Oxtr+/-73. В этих исследованиях тестировались взрослые мыши. В очень недавнем исследовании74 Nunes, Gliksberg и коллеги наблюдали нарушение социальной привязанности у взрослых данио-рерио после химической абляции окситоцинергических нейронов в дорсоростральной части нейросекреторной преоптической области на ранних стадиях развития (4–6 dpf и 12–14 dpf), но более поздняя абляция (20–22 dpf и 90 dpf) не повлияла на социальную предпочтительность взрослых рыб. Поскольку абляционные окситоцинергические нейроны полностью восстанавливались в течение 42 дней74, описанная нарушенная социальная предпочтительность, вероятно, связана с необходимостью этих окситоцинергических нейронов на ранних стадиях развития. Нокаутные рыбы, которые мы использовали в этом исследовании, имели недостаток Oxtr и Oxtrl соответственно на всех стадиях развития. Для oxtr−/− мы наблюдали значительное снижение социальной предпочтительности между пиком максимальной социальной предпочтительности в 3 wpf и 8 wpf. Возможно, это значительное снижение сохраняется до взрослого состояния, что приводит к нарушенной социальной предпочтительности у взрослых oxtr−/−, но это еще предстоит проверить с использованием нокаутных рыб. По сравнению с рыбами oxtrl−/−, поддержание социальной предпочтительности было нарушено в большей степени у рыб oxtr−/−, что предполагает, что этот рецептор может быть более важен для развития взрослого аффилиативного поведения, опосредованного окситоцином, на личиночных стадиях.
Наши данные не указывают на изменение уровня тревожности или агрессии у рыб oxtr−/− и oxtrl−/− в тестируемом возрасте, поскольку два показателя тревожности — смелость (см. Дополнительный Рис. S2c) и замирание (см. Дополнительный Рис. S3c и S3d) — не отличались между дикими и мутантными рыбами. Кроме того, частота агрессивного поведения, такого как укусы, не изменилась. В соответствии с исследованием Ribeiro и др.53, исследование с нокаутом рецептора окситоцина у луговых полевок75 показало нарушение социального распознавания у Oxtr−/−, но не выявило значительных изменений в социальном поведении. Наши результаты также показывают, что Oxtr и Oxtrl не имеют одинаковых функций, поскольку нокауты oxtr−/− и oxtrl−/− демонстрируют разную степень социальной предпочтительности на протяжении развития. Важно отметить, что при рассмотрении роли рецепторов окситоцина может потребоваться оценка потенциальной роли вазотоцина, ортолога млекопитающего вазопрессина у данио-рерио, который также может связываться с Oxtr и Oxtrl, хотя и с гораздо меньшей аффинностью, чем Oxt52.
Поскольку рыбы oxtrl−/− демонстрируют усиленный уровень социальной предпочтительности в 3 и 4 wpf, мы ожидали, что они будут стайться более плотно, с уменьшенным расстоянием между группами по сравнению с дикими рыбами. Однако увеличение социальной предпочтительности не сопровождалось изменениями в параметрах стайного поведения — расстоянии до ближайшего соседа, межвидовом расстоянии, расстоянии до самого дальнего соседа и параметре поляризации «объясненная дисперсия» ни у oxtr−/−, ни у oxtrl−/− в 4 wpf. И все же, в 8 wpf расстояние до ближайшего соседа, межвидовое расстояние и расстояние до самого дальнего соседа были значительно увеличены, а объясненная дисперсия была значительно снижена как у oxtr−/−, так и у oxtrl−/−. Эти данные указывают на менее поляризованное стайное поведение с увеличенным расстоянием между группами у oxtr−/− и oxtrl−/−. В соответствии с нашими результатами, интраперитонеальное введение L368,899 также привело к увеличению расстояния до ближайшего соседа, межвидового расстояния и расстояния до самого дальнего соседа в стаях из четырех взрослых рыб52, подтверждая про-сплоченную роль двух рецепторов окситоцина в организации стай у рыб старше 8 wpf. Основные различия между экспериментами по изучению стайного поведения Landin и коллег и нашими — это возраст (взрослые против 8 или 4 wpf), размер стаи (4 рыбы против 20 рыб в стае) и поведенческая камера (трапециевидная, 1,8 л против круглой, 2,4 или 3,0 л). Кроме того, в исследовании Landin и др. оба рецептора окситоцина были заблокированы введением антагониста за 1 час до эксперимента, тогда как в наших экспериментах только один из двух рецепторов окситоцина был нокаутирован с момента оплодотворения. Хотя и не было значительным, первые признаки сниженной способности к стайному поведению у oxtrl−/− были заметны уже в возрасте 4 недель. Поскольку стайное поведение непрерывно развивается до взрослого состояния17, наиболее правдоподобным объяснением для обоих наборов данных является то, что рецепторы окситоцина, вероятно, важны для организации стаи на более поздних стадиях развития, в то время как другие сигнальные пути могут компенсировать в 4 wpf. Подобно социальной предпочтительности, оба рецептора окситоцина модулируют стайное поведение, но в разной степени. Поскольку на параметры стайного поведения также влияют избегание хищников76,77 и опыт78, стайное поведение не следует рассматривать как исключительное проявление социального поведения. Кроме того, стайное поведение регулируется как зрительными, так и механосенсорными (боковая линия)79 входами.
В исследовании Tang и коллег80 сравнивалось стайное поведение 90 нокаут-линий CRISPR/Cas9 (по разным генам) (6 взрослых рыб в стае). Это исследование выявило влияние множества генов на коллективное поведение данио-рерио, в частности, на скорость плавания, расстояние между группами и поляризацию. Tang и др. также показали, что один ген не обязательно влияет на скорость плавания, расстояние между группами и поляризацию, а может влиять только на один или два из этих параметров стайного поведения. Наши данные согласуются с этим выводом, поскольку мы не обнаружили значительных изменений в суммарном расстоянии стаи в 8 wpf (см. Дополнительный Рис. S2b), тогда как параметры расстояния между группами и поляризации были значительно изменены в этом возрасте. На первый взгляд, наблюдения, что рыбы с мутацией рецептора окситоцина преждевременно развивают предпочтение к социальным компаньонам, но демонстрируют плохую координацию и стайное поведение в 8 wpf, могут показаться противоречивыми. Однако это может быть объяснено наблюдениями снижения предпочтения к социальным компаньонам в онтогенезе. Мутанты oxtr−/−, например, демонстрируют больший эффект в снижении параметров стайного поведения в 8 wpf по сравнению с oxtrl−/−, что согласуется с большим снижением предпочтения к социальным компаньонам в этом возрасте. Незначительные или отсутствующие изменения в параметрах стайного поведения в 4 wpf могут отражать либо технические ограничения этого анализа на раннем возрасте, либо наличие «потолочного эффекта» для обнаружения улучшенных характеристик стайного поведения.
В совокупности наши результаты показывают, что рецепторы окситоцина играют важную роль в развитии и поддержании социального поведения данио-рерио, и влияние окситоциновой сигнализации зависит от возраста и среды обитания рыб.
Материалы и методы
Раздел «Материалы и методы» соответствует рекомендациям ARRIVE 2.081.
Дизайн исследования, размер выборки и критерии исключения
Количество биологических репликатов для каждого эксперимента можно найти в Дополнительной таблице S1. Для исследования социальной предпочтительности мы планировали протестировать 18 рыб на группу. Размер выборки определялся до экспериментов с использованием метода E‑equation82. К сожалению, некоторые экспериментальные рыбы погибли во время содержания, поэтому на группу тестировалось от 15 до 18 рыб. Данные исключались из дальнейшего анализа, если средняя скорость плавания тестовой рыбы была ниже порогового значения (73/411). Этот порог рассчитывался для каждой возрастной группы путем определения средней скорости плавания каждой генотипической группы (дикий тип, oxtr−/− или oxtrl−/−) и умножения наименьшей на 0,6. Рыбы трех генотипических групп (oxtr+/+;oxtrl+/+ = «дикий тип», oxtr−/−;oxtrl+/+ = «oxtr−/−» и oxtr+/+;oxtrl−/− = «oxtrl−/−») содержались либо в изоляции, либо с сородичами при плотности 10–15 рыб/л и тестировались на социальную предпочтительность в возрасте 2, 3, 4 или 8 wpf. Чтобы условия тестирования оставались максимально схожими, разные генотипы тестировались без излишней задержки (все эксперименты одной возрастной группы проводились в течение одного месяца, за исключением 8 wpf (группы тестировались в течение четырех месяцев), а рыбы, выращенные в изоляции и социальных условиях, одной генотипической группы тестировались в один день, когда это было возможно).
В 30-минутном эксперименте по стайному поведению параметры стайного поведения анализировались в стае из 20 рыб в возрасте 4 или 8 wpf в трех различных генотипах (oxtr+/+;oxtrl+/+ = «oxtr+/+» или «oxtrl+/+», oxtr−/−;oxtrl+/+ = «oxtr−/−» и oxtr+/+;oxtrl−/− = «oxtrl−/−»). В отличие от социальной предпочтительности, мы обнаружили значительную разницу в некоторых параметрах стайного поведения между гомозиготными дикими кузенами oxtr−/− и гомозиготными дикими кузенами oxtrl−/−. Поэтому мы анализировали диких данио-рерио как две генотипические группы: «oxtr+/+» и «oxtrl+/+» соответственно. Мы протестировали 23–28 стай, выращенных в социальных условиях (10–15 рыб/л), на группу.
Тестовые рыбы использовались только один раз, после чего их умерщвляли и генотипировали. Если рыба/стая идентифицировалась как гетерозиготная (22/611), она исключалась из дальнейшего анализа.
Экспериментальные животные и получение нокаутов
Гены oxtr (ZDBGENE1108052/NCBI mRNA Reference Sequence: NM_001199370.1) и oxtrl (ZDBGENE1108051/NCBI mRNA Reference Sequence: NM_001199369.1) были мутированы Ajay Mathuru и Caroline Kibat с использованием системы sgRNA:Cas9, описанной в83. Два CRISPR-таргета (GGAAGTTACCGTGTTGGCCT и GGCTGATAAGCTTTAAAATA для oxtr; GTGCGTCCTTGTGGCCATCC и GGGGGGATTTTGTTCAGCCC для oxtrl) для каждого гена были идентифицированы с помощью ZiFit (http://zifit.partners.org/zifit/). Специализированные sgRNA с 20-нуклеотидной последовательностью, комплементарной целевому участку, были синтезированы путем клонирования целевых последовательностей в экспрессионный конструкт pDR274 (Addgene #42,250), а затем in vitro транскрибированы с использованием T7-промотора согласно протоколу производителя (Thermo Fisher # AMB13345).
мРНК Cas9 была транскрибирована из линеаризованной плазмиды MLM3616 (Addgene #42,251) согласно протоколу производителя (Thermo Fisher # AMB1344). Коктейль sgRNA:Cas9 RNAs (содержащий 12,5 нг/мкл sgRNA и 300 нг/мкл Cas9) вводили в одноклеточные эмбрионы дикого типа AB. Эффективность CRISPR-таргетов и качество эндонуклеазы Cas9 определяли через 24 часа после введения sgRNA:Cas9 путем ПЦР на 10% инъецированных эмбрионов. Оставшиеся эмбрионы выращивали до взрослого состояния и генотипировали через три месяца после инъекции. Всего было генотипировано 64 особи. Последовательность праймерной пары для генотипирования подробно описана в разделе «Методы генотипирования». ПЦР-продукты были клонированы в pGEM-T (Promega #1360) и секвенированы для проверки мутаций. F0 мутанты затем скрещивались с Danio Reds (https://doi.org/10.1016/S0006291X(03)012828). Рыбы F1 инкроссировались для создания гомозиготных мутантных линий, передающих зародышевую линию.
Линии названы oxtrync02 и oxtrlync03, в данной работе сокращенно oxtr−/− и oxtrl−/− соответственно. Гомозиготные дикие кузены этих рыб использовались в качестве контроля дикого типа. Кроме того, эти дикие кузены использовались для регулярного ауткроссинга гомозиготных нокаутов (KO) для максимально возможного сохранения генетического фона экспериментальных рыб. Мы не смогли протестировать двойные KO рыбы oxtr−/− oxtrl−/−, поскольку их получение путем скрещивания не удалось, возможно, из-за того, что присутствие по крайней мере одного рецептора окситоцина жизненно важно на определенных стадиях развития. При оценке экспрессии нокаутированного рецептора с использованием специфических праймеров с помощью ОТ-ПЦР, амплификационные сигналы обнаруживались у дикого типа, но не у мутантных рыб, что указывает на успешный нокаут. Более того, компенсация нокаутированного рецептора его ортологом, рецептором окситоцина, не была обнаружена (см. Дополнительный Рис. S1b).
Экспериментальные рыбы разводились путем инкроссинга гомозиготных мутантных или диких рыб в соотношении 1:1 по полу на партию. Каждая группа состояла из экспериментальных рыб из нескольких кладок (до 17), а размер кладки составлял 300–400 яиц (без различий между дикими и мутантными рыбами). В день эксперимента тестовым рыбам было 2, 3, 4 или 8 wpf, в зависимости от возрастной группы. Мы использовали экспериментальных рыб с общей длиной тела, сравнимой с общей длиной тела, описанной в «Zebrafish Book»66: 6 мм (2 wpf), 8 мм (3 wpf), 10 мм (4 wpf) и 14 мм (8 wpf). Различий в развитии или росте, сравнивая мутантных и диких рыб, не выявлено. В тесте социальной предпочтительности стимулирующие рыбы (две на тестовую рыбу) имели тот же возраст и схожий размер, что и тестовые рыбы. Поскольку отличить самцов и самок на стадиях развития, используемых в этом исследовании, трудно (ювенильные) или невозможно (личинки), рыбы становились экспериментальными независимо от пола.
Содержание и уход за данио-рерио
До 5 dpf личинки содержались в инкубаторе при 28,5 °C с 14-часовым световым/10-часовым темным циклом. В 2 dpf яйца либо индивидуально изолировались в 12-луночной планшете (3 мл среды E3 [5 мМ NaCl, 17 мМ KCl, 0,33 мМ CaCl2, 0,33 мМ MgSO4] на лунку диаметром 22 мм), либо содержались в чашках Петри (диаметром 145 мм, заполненных 150 мл среды E3) группами по 50 штук.
С 5 dpf они выращивались в изоляции (включая визуальные барьеры, размещенные в каждом втором резервуаре, см. Рис. 2а, центр) в резервуарах ZebTEC объемом 1,1 л или в группах со смешанным полом при плотности 10–15 рыб/л в резервуарах ZebTEC объемом 1,1 л (тест социальной предпочтительности) или 3,5 л (тест стайного поведения).
С 5 dpf данио-рерио содержались в системе ZebTEC Active Blue Stand Alone System при температуре воды 28,5 °C ± 1 °C, pH = 7,4 ± 0,3 и проводимости = 650 µS ± 100. Цикл освещения/темноты в виварии составляет 14 часов света / 10 часов темноты с 30-минутной фазой сумерек. Рыбы кормились три раза в день: до 10 dpf — нематодами (Turbatrix aceti, культивируемыми в лаборатории), CAVIAR 50–100 (SAFE) и измельченным Gemma Micro 75 (Skretting); с 10 до 20 dpf — нематодами, артемией (Artemia salina, Ocean nutrition V154019) и CAVIAR 50–100; с 21 до 56 dpf — артемией и CAVIAR 100–200 (SAFE).
Экспериментальные процедуры
Тест социальной предпочтительности
Поведенческая камера (высотой 10 мм) для тестирования социальной предпочтительности состояла из тестовой зоны (25 × 75 мм (для 2–4 wpf) и 50 × 75 мм (для 8 wpf)) и зон стимуляции (8 × 75 мм (для 2–4 wpf) и 16 × 75 мм (для 8 wpf)). Поведенческая камера помещалась на экран, обеспечивая белую фоновую подсветку, а камера, расположенная сверху, записывала движения рыбы с частотой 20 (3–8 wpf) или 30 (2 wpf) кадров в секунду с помощью программы «Pylon recorder». Для уменьшения визуальных и акустических помех установка окружалась черной звукопоглощающей коробкой (ДШВ: 450 × 670 × 850 мм). Перед экспериментом тестовая и стимулирующая рыба перемещалась из своего домашнего резервуара в 1-литровый разводной резервуар (TECNIPLAST, артикул: ZB10BTE) с вставкой для выведения мальков (TECNIPLAST, артикул: ZB300BTI) и содержалась в инкубаторе при 28,5 °C (тестовая и стимулирующая рыба в разных резервуарах). После каждого теста поведенческая камера очищалась горячей водой (~60 °C) и заполнялась свежей водой из системы ZebTEC Stand Alone (28,5 °C). Рыб перемещали с помощью одноразовой пипетки объемом 3 мл (конец был отрезан для получения достаточного диаметра), поднимая вставку для выведения мальков, а затем помещали в центр тестовой зоны (тестовая рыба) или на край зоны стимуляции (стимулирующая рыба) поведенческой камеры. В 8 wpf камера закрывалась крышкой от чашки диаметром 145 мм, чтобы предотвратить выпрыгивание рыбы. Тестовая рыба адаптировалась к камере без других рыб в течение десяти минут, за которыми следовала 12-минутная тестовая фаза, в течение которой две стимулирующие рыбы того же возраста и размера помещались в одну из зон стимуляции. Прозрачная стенка между тестовой и стимулирующей зоной обеспечивала визуальный доступ к стимулирующим рыбам, в то время как белая непрозрачная стенка определяла заднюю сторону стимулирующей зоны.
Эксперимент по стайному поведению
Установка для изучения стайного поведения (см. Дополнительный Рис. S2a) состояла из белой круглой поведенческой камеры диаметром 70 см, окруженной водой температурой 29,5 °C (подогреваемой насосом) для предотвращения охлаждения воды внутри поведенческой камеры во время 30-минутного эксперимента. Высокоразрешающая камера (Basler acA4112-30um), расположенная на высоте 73 см над камерой, записывала рыбу с частотой 30 кадров в секунду с помощью программы «Pylon recorder», а 1200 белых светодиодов на потолке обеспечивали необходимое освещение (примерно 650 люкс). Белая коробка, окружающая установку, снижала визуальные и акустические помехи для рыб. Для каждого повторения камера полностью опорожнялась, высушивалась и заполнялась 2,4 л (4 wpf) / 3,0 л (8 wpf) свежей теплой (28,5 °C) водой из системы Stand Alone. Утром в день эксперимента домашние резервуары перемещались из системы ZebTEC Stand Alone в инкубатор при 28,5 °C. Примерно за 5 минут до начала эксперимента 20 рыб одного возраста и схожего размера переносились из их домашнего резервуара в 1-литровый разводной контейнер (TECNIPLAST) с вставкой для выведения мальков (TECNIPLAST, артикул: ZB300BTI). С помощью вставки для выведения мальков все 20 рыб одной стаи одновременно перемещались в центр поведенческой камеры. Запись стайного поведения начиналась немедленно и продолжалась в течение 30 минут.
Генотипирование
Для идентификации гомозиготных нокаутов (KO) и гомозиготных диких типов (wt) при создании экспериментальных рыб выполнялась ПЦР с использованием выщипов плавников в соответствии с протоколами генотипирования ZIRC (https://zebrafish.org/wiki/protocols/genotyping). Дополнительно каждая экспериментальная рыба генотипировалась после эксперимента. Использовались следующие последовательности праймеров:
| Прямой | Обратный | Размер полосы | |
|---|---|---|---|
| oxtr | TGGAGGACATCTTCAAGGACTT | CCCCTCGGTGCTTCAGGA | 916 п (wt), 413 п (KO) |
| oxtrl | TGATCCTCTGGCCCATTAACT | GGTCCATAAAAGCGAAAGG | 314 п (wt), 304 п (KO) |
ПЦР-продукты загружались на 1% (oxtr) или 2% (oxtrl) агарозный гель и прогонялись при 100 В в течение 30 минут (oxtr) или > 2 часов (oxtrl). Но поскольку разница между oxtrl+/+ и oxtrl−/− составляет всего 10 пар оснований, ПЦР-продукты также секвенировались с использованием прямого праймера.
Реалтайм ПЦР
РНК экстрагировалась из изолированных мозгов, как описано ранее67, и с использованием QuantiTect Reverse Transcription Kit (QIAGEN Cat. No. 205311) 200 нг РНК транскрибировались в кДНК. 5 мкл 1:10 разведенной кДНК-матрицы смешивались с 1,3 мкл праймеров (10 мкМ) и 6,25 мкл SYBR Green PCR master mix (Applied Biosystems/Thermo Fisher Art. No. 4309155). Параметры циклов составляли 10 минут при 95 °C, затем 40 циклов денатурации (15 с при 95 °C) и амплификации (60 с при 60 °C). Для qRTPCR использовалась система Real Time PCR System (Applied Biosystems/Thermo Fisher Art. No. 4376600). Порог флуоресценции устанавливался на 0,9 для всех экспериментов и генов, и мы использовали TATA-связывающий белок (tbp) в качестве референсного гена. Мы использовали метод ΔCT и следующие последовательности праймеров:
| Прямой | Обратный | |
|---|---|---|
| oxt | AAGCTCTCGGTGTCAGCCTTG | GCATACACTGTCGAATGGG |
| oxtr | CCAACCTCTTCGTTCTTTACAC | AAAATCTCCCCAGCAATC |
| oxtrl | AAACCTGTGCGTCCTTGTGAT | AAAAGCGAAAGGTGATGTCC |
| tbp | GTACTCACAGGTGCCAAGGTG | ATTGCGTAGGTCACCCCAG |
Измеряемые параметры
В тесте социальной предпочтительности положение тестовой рыбы измерялось в каждом кадре акклиматизации и в течение последних 10 минут тестовой фазы, поскольку стимулирующим рыбам требовалось до 2 минут после перемещения в зону стимуляции для проявления нормального социального поведения. Во время анализа тестовая зона была разделена на две области интереса (ROI). «Социальная ROI» — это область рядом со стимулирующими рыбами, а «антисоциальная ROI» — напротив тестовой зоны. Процент общего времени, в течение которого рыба находилась в «социальной ROI», определялся. Средняя скорость плавания в отсутствие (акклиматизация) или присутствии (тест) стимулирующих рыб рассчитывалась и использовалась в качестве критерия исключения для рыб с пониженной подвижностью (например, из-за замирания).
В экспериментах по стайному поведению все двадцать рыб каждой стаи отслеживались с помощью TRex84 (порог отслеживания 15 (4 wpf) и 50 (8 wpf)), и рассчитывались расстояние до ближайшего соседа, межвидовое расстояние и расстояние до самого дальнего соседа каждой рыбы, а также суммарное расстояние стаи и объясненная дисперсия как параметр скоординированного плавания. Для определения объясненной дисперсии компоненты x и y всех индивидуальных траекторий (20 на стаю) сначала собирались в матрицу 40 × 54 000 (30 минут при 30 кадрах в секунду), ковариационная матрица которой затем подвергалась eigen-разложению для получения главных компонент (PC). Мы обнаружили, что первые две PC соответствовали компонентам x и y траектории центра стаи во времени. Анализ главных компонент (PCA) — это подход к снижению размерности, который может быть использован для количественной оценки степени линейной корреляции в наборе данных. Если отдельные признаки сильно коррелируют, их можно сжать в одну PC, практически не теряя данных. Это можно измерить дисперсией исходного набора данных, которая объясняется полученными PC. В нашем случае, чем выше дисперсия, объясненная первыми двумя PC, тем выше корреляция между движением каждой рыбы и движением стаи в целом (см. Дополнительный Рис. S2d). Дополнительно мы анализировали объясненную дисперсию у 20 рыб из случайно выбранных видео, ожидая отсутствия скоординированного плавания. В соответствии с этим ожиданием объясненная дисперсия была очень низкой и непрерывно увеличивалась с увеличением числа рыб из одного видео (см. Дополнительный Рис. S2e). Таким образом, объясненная дисперсия может быть использована для количественной оценки скоординированного плавания рыб.
Статистические методы
Значимость указывается следующим образом: p < 0,05, p < 0,01, p < 0,001. Все значения значимости можно найти в Дополнительной таблице S2.
Нормальность распределения данных проверялась с помощью теста Колмогорова-Смирнова, который выявил ненормальное распределение. Для несвязанных данных использовался тест Краскела-Уоллиса с последующим апостериорным тестом Манна-Уитни U (= тест ранговых сумм Уилкоксона). Статистический анализ проводился с использованием Python (из scipy.stats: kstest, kruskal и mannwhitneyu) или MATLAB (kstest, signrank, kruskalwallis и ranksum). В тесте социальной предпочтительности в анализ было включено n = 332 рыбы (79 исключено), в эксперименте по стайному поведению в анализ было включено n = 184 стаи (18 исключено).
Ослепление и рандомизация
Исследователи не были ослеплены по отношению к генотипу экспериментальных рыб, но оба эксперимента записывались с помощью верхней камеры и анализировались с использованием программного обеспечения для отслеживания (пользовательский скрипт для отслеживания и анализа социальной предпочтительности и TRex84 плюс пользовательский скрипт Python для анализа стайного поведения) для обеспечения объективного анализа данных. Тестовые рыбы случайным образом выбирались из их домашней группы, а в тесте социальной предпочтительности расположение стимулирующих рыб между повторениями менялось в случайном порядке. Эксперименты проводились с 8:00 до 17:00, и порядок генотипических групп в день менялся между экспериментальными днями, чтобы минимизировать потенциальное влияние циркадных ритмов. Кроме того, положение резервуара во время содержания выбиралось таким образом, чтобы все повторения одной экспериментальной группы содержались во всех возможных высотах/условиях освещения.
Заявление об этике
Все процедуры проводились в соответствии с ведомственными руководящими принципами Общества Макса Планка и были одобрены Регенсбушпрезидиумом Дармштадта, Германия (государственный идентификатор: V 5419 c20/15F126/1013 и V5419 c20/15F126/1016).