Интересное сегодня
Стратегии для разрешения сложных психоделических переживаний...
Введение Психоделические вещества, такие как псилоцибин и N,N-диметилтриптамин (ДМТ), имеют долгую и...
Эффективность программ поддержки психического здоровья для с...
Профессиональный стресс у сотрудников исправительных учрежденийСотрудники исправительных учреждений ...
Дедиктивный тематический анализ: руководство по применению и...
Может ли тематический анализ быть дедуктивным? Да, тематический анализ может быть дедуктивным. Это г...
Как справиться с гневом ребенка: простая фраза для родителей
Введение Гнев является одной из самых сложных эмоций, с которой детям нелегко справляться. Для родит...
Взаимодействие человека и робота: чувство собственной активн...
Введение: Понимание чувства собственной активности (SoA) Чувство собственной активности (Sense of A...
Как формируется идентичность в пожилом возрасте: исследовани...
Развитие идентичности в пожилом возрасте Идентичность — это наше ощущение непрерывности «Я», стабиль...

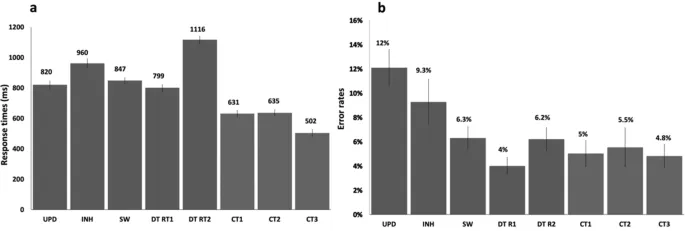
Исследование единства и разнообразия исполнительных функций в приборс исследовании с использованием фМРТ
Введение
Система центрального исполнительного контроля (СЦИК), являясь ключевым компонентом рабочей памяти, регулирует мыслительные процессы и поведение человека путем поддержания и манипулирования информацией в системах хранения данных. Было высказано предположение, что СЦИК может быть разделена на отдельные исполнительные подфункции, такие как ингибирование, переключение и обновление. Вопрос о том, являются ли эти исполнительные функции (ИФ) отличными друг от друга (разнообразие) или они опираются на общий механизм или ресурс (единство), остается предметом активных исследований. Несмотря на различные точки зрения, предыдущие поведенческие исследования в целом предполагали, что речь идет о комбинации обоих аспектов, особенно у взрослых. Например, Мияке и коллеги исследовали единство и разнообразие исполнительных функций, используя различные тесты ИФ в поведенческом эксперименте. Они выделили три основных фактора: обновление (обновление и мониторинг ментальных репрезентаций), ингибирование (подавление нерелевантных для задачи стимулов, которые могут вызывать помехи) и переключение (также называемое сдвигом; гибкость в переключении внимания между двумя задачами, операциями или ментальными установками). Эти факторы коррелировали друг с другом, что подтверждает идею единства, но также имели уникальную вариативность, поддерживающую идею разнообразия. Поскольку выполнение двойной задачи (одновременное выполнение двух задач) не коррелировало с этими ИФ, было отмечено, что выполнение двойной задачи может составлять отдельную ИФ. Следуя трехфакторной модели Мияке и коллег, выполнение двойной задачи в основном игнорировалось, и некоторые исследования предполагали, что эти три ИФ могут быть связаны с общим высшим когнитивным контролем. Наконец, Фридман и коллеги предложили вложенную модель, предполагающую, что обновление и переключение лежат в основе единства и разнообразия ИФ, в то время как ингибирование не имеет специфического измерения, поскольку оно рассматривается только как часть общей вариативности.
Эти выводы поведенческих исследований поднимают вопрос о функциональных нейроанатомических коррелятах исполнительных функций. Активируют ли различные ИФ, такие как обновление, ингибирование, переключение и выполнение двойной задачи, одни и те же или разные области мозга? Насколько нам известно, этот важный вопрос исследовался лишь фрагментарно, и результаты были довольно противоречивыми. Например, некоторые исследования использовали межсубъектные дизайны (то есть разные выборки участников для каждой ИФ), что затрудняло прямое сравнение областей мозга, вовлеченных в различные ИФ. Одно эмпирическое исследование, проведенное Коллетт и коллегами, оценивало три исполнительные функции (обновление, переключение и ингибирование) во время ПЭТ-сканирования. Результаты показали области единства в теменных областях (то есть правый внутритеменной борозда и левая верхняя теменная извилина) и области разнообразия во фронтотеменных областях. Хотя предполагается, что ИФ обслуживаются фронтотеменной сетью, отсутствие областей единства в лобных долях показалось несколько удивительным. Потенциальным ограничением этого исследования могли быть различные выборки для тестирования ИФ (межсубъектный дизайн) и задачи, которые были довольно разными по своей природе. Подробно, индивидуальные различия, сопровождаемые различными стратегиями выполнения задач, могут повлиять на однородность функций.
Чтобы обойти проблемы, связанные с межсубъектными дизайнами, некоторые исследования использовали внутрисубъектные дизайны, то есть каждый участник выполнял несколько задач ИФ. Однако предыдущие исследования обычно ограничивались изучением только двух ИФ одновременно. В соответствии с приведенными выше выводами, ряд исследований связывал ИФ в целом с фронтотеменной сетью, однако другие исследования предполагали отдельные локализации для этих функций. Такие расходящиеся результаты наблюдались не только для разных ИФ, но и для одной и той же функции. Например, при исследовании ингибирования и переключения Хедден и Габриэли наблюдали доказательства единства (то есть обе ИФ активировали одни и те же области), в то время как Сильвестр и коллеги нашли доказательства не только единства, но и разнообразия (то есть области, активированные только одной ИФ). Потенциальным объяснением этого расхождения могут быть различия в статистической мощности исследований или то, что оба исследования использовали очень разные парадигмы задач. Следовательно, настоящее исследование использовало высокосравнимые задачи для оценки четырех ИФ, которые были выведены из одной и той же базовой парадигмы, используя один и тот же набор стимулов. Взятые вместе, исследования, изучающие две ИФ одновременно, в значительной степени поддерживают идею о том, что нейронные корреляты ИФ могут демонстрировать перекрытие (единство). Однако они по-прежнему ограничены тем, что изучают только две ИФ одновременно и дают частично противоречивые результаты.
Мы также рассмотрели исследования, посвященные функциональным нейроанатомическим коррелятам только одной ИФ. Эти исследования использовали связанные задачи для целевой ИФ (например, n-назад для обновления, чередование задач для переключения, задачи Струпа для ингибирования и комбинация двух простых задач для двойной задачи) и в целом демонстрировали фронтотеменную сеть для каждой ИФ, но также предполагали некоторые ключевые области для точных требований задачи. Ключевой областью для ингибирования может быть нижняя лобная извилина (НЛИ) (BA 46) для разрешения конфликтов, для обновления ключевой областью может быть надкраевая извилина (НКИ), как один из основных каналов связи между лобной и теменной областями, действующий как врата, облегчающие обновление информации в рабочей памяти, для выполнения двойной задачи области, такие как средняя лобная извилина (СЛИ) и внутритеменная борозда (ВБ), могут особенно участвовать в контроле порядка задач. Помимо областей единства, активация таких областей в одной или нескольких задачах в разной степени может указывать на то, что естественный процесс каждой ИФ может составлять подпроцессы в дополнение к их уникальным требованиям. Например, обновление включает кодирование и избирательное поддержание информации в рабочей памяти. Этот процесс в некоторой степени включает ингибиторный процесс, поскольку устаревшая информация должна подавляться для обработки информации. Аналогично, выполнение двойной задачи может в некоторой степени включать другие ИФ, поскольку необходимо подавлять вторую задачу до тех пор, пока первая задача не будет обработана, а затем переключиться на вторую задачу. Взятые вместе, активация в определенных областях может быть объяснена парными корреляциями только двух ИФ, а некоторая вариативность была уникальной для каждой ИФ.
Наконец, метаанализы и обзорные исследования, изучающие единство и разнообразие, в основном ограничивались двумя или тремя ИФ, затрагивая заднюю часть лобных областей и теменные области для единства и различные области фронтотеменных областей для разнообразия. Например, Деррфус и коллеги предложили нижнюю фронтальную границу (НФГ), расположенную в задней части верхней лобной коры, которая играет важную роль в обработке как переключения, так и ингибирования. Другое метааналитическое исследование изучало общие и различные области, связанные с выполнением двойной задачи и переключением, затрагивая двустороннюю нижнюю теменную борозду (НТБ) и верхнюю теменную долю (ВТД) для единства в дополнение к различным областям для каждой функции во фронтотеменных областях. Активация НЛИ и ВТД для единства ИФ также была отмечена в обзоре Коллетт и коллег. Эти исследования продемонстрировали, что функциональные нейроанатомические корреляты ИФ могут отражать закономерность, найденную для когнитивных представлений этих ИФ, а именно, что все ИФ могут активировать некоторые общие перекрывающиеся области (единство), которые включали теменные и задние верхние лобные доли, такие как левая нижняя фронтальная борозда (НФБ), нижняя прецентральная борозда (НПБ) и левая ВТД, но также каждая активирует свои уникальные отдельные области (разнообразие), такие как правая НЛИ для ингибирования. Однако точное расположение нейронных коррелятов конкретных ИФ может зависеть от конкретной используемой парадигмы и может различаться, по крайней мере, в некоторой степени, между выборками. Следовательно, остается открытым вопрос, является ли текущая закономерность единства и разнообразия простым артефактом сравнения между различными исследованиями или же она отражает истинные функциональные нейроанатомические корреляты ИФ.
Взятые вместе, предыдущие данные о единстве и разнообразии нейронных коррелятов различных ИФ ограничены исследованиями, изучающими только одну ИФ, исследованиями, использующими межсубъектные дизайны, или исследованиями с внутрисубъектными дизайнами, обычно изучающими не более двух ИФ одновременно. Следовательно, целью данного исследования было оценить функциональные нейроанатомические корреляты четырех исполнительных функций: обновления (задача n-назад), ингибирования (задача Струпа), переключения (задача переключения задач) и выполнения двойной задачи (задача PRP) в внутрисубъектном дизайне. Чтобы минимизировать влияние задач, мы создали четыре различных версии, основанные на одной и той же базовой задаче, которые основаны на одних и тех же стимулах и в основном отличаются инструкциями, данными участникам. В дополнение к этим ключевым задачам, мы также создали соответствующие контрольные задачи, которые разработаны для соответствия перцептивным и моторным требованиям и отличаются только по своим требованиям к соответствующей ИФ, так что могут быть идентифицированы специфические для ИФ активации. В деталях, ИФ обновления определялась сравнением задачи 1-назад с задачей 0-назад, ИФ ингибирования сравнением несогласованных и согласованных стимулов Струпа, ИФ переключения сравнением условия переключения задач с условием повторения задач, и, наконец, ИФ выполнения двойной задачи сравнением выполнения двойной задачи с выполнением одной задачи (см. «Методы» для деталей). В отношении анализа мы будем использовать конъюнктивный анализ для проверки перекрытия четырех паттернов активации, специфичных для ИФ (единство), и парные сравнения между активациями, специфичными для ИФ, для проверки разнообразия. Эти анализы будут подкреплены детальными анализами бета-значений, полученных из индивидуальных пиковых координат этих контрастов.
Методы
Участники
В эксперименте приняли участие 29 участников в возрасте от 18 до 30 лет (14 мужчин: средний возраст 23,50 лет, стандартное отклонение 3,32, и 15 женщин: средний возраст 23,50 лет, стандартное отклонение 3,32). Каждый участник дал письменное информированное согласие и получил £30 за участие. Мы использовали следующие критерии исключения: наличие любого прошлого или текущего серьезного медицинского, неврологического или психиатрического заболевания, которое могло бы снизить когнитивные функции; прием психоактивных препаратов; употребление алкоголя; употребление ≥ 8 чашек или ≥ 900 мг кофеина; более 15 баллов по шкале депрессии Бека; дальтонизм. Исследование было одобрено этическим комитетом Департамента наук о жизни Университета Брунеля в Лондоне. Эксперимент проводился в соответствии с соответствующими руководящими принципами и нормами этического комитета Департамента наук о жизни Университета Брунеля в Лондоне и Хельсинкской декларацией.
Дизайн задачи и процедура
Участники лежали на спине в МРТ-сканере, держа в руках две МРТ-совместимые панели ответа, в МРТ-совместимых внутриканальных наушниках и смотрели на экран через систему зеркал. Мы использовали блочный дизайн для проведения эксперимента. Было четыре основных условия задачи (ИФ задачи), и каждый блок этих условий состоял из 10 испытаний: обновление, ингибирование, переключение и выполнение двойной задачи. В дополнение к этому, мы добавили три условия одиночной задачи (контрольные задачи), каждый блок состоял из шести испытаний, и период отдыха (базовый уровень) для создания подходящих контрастов. Продолжительность блоков варьировалась в зависимости от условия: блоки ИФ длились 25 с, блоки контрольных задач длились 15 с, а базовый отдых длился 15 с (см. также раздел «Анализ данных»). Блоки предъявлялись случайным образом с десятью повторениями каждого блока. Эксперимент начинался с 15-секундного блока базового уровня (состоящего только из фиксационного креста), за которым следовали семь исполнительных и контрольных условий, предъявлявшихся случайным образом в 10 циклах. Эксперимент завершался еще одним 15-секундным блоком базового уровня. Каждый блок предварялся инструкцией, отображаемой в течение 4,5 с, за которой следовало переменное количество испытаний (см. ниже), каждое длительностью 2,5 с. Все испытания состояли из 250 мс фиксации, 500 мс стимула, 2000 мс интервала ответа (начиная с момента появления стимула) и, наконец, 250 мс фазы обратной связи, которая представляла «X» при ошибке и пустой экран в противном случае.
Мы использовали цифры (1, 2) и цветовые слова (Красный, Зеленый, Желтый, Синий), которые предъявлялись на экране с белым фоном либо по отдельности, либо в комбинации одной цифры и одного слова (например, 1 Красный). Слова могли быть либо в согласующемся цвете (слово «Красный» красным цветом), либо в несогласующемся цвете (слово «Красный» зеленым цветом), либо в нейтральном цвете (слово «Красный» черным цветом). Участники всегда отвечали на цифры левой рукой, а на слова — правой рукой. На основе этого мы создали различные задачи путем небольших вариаций в отображаемых стимулах и инструкциях по ответу на них.
Контрольные задачи (КЗ)
Для первой контрольной задачи (КЗ1) целевым стимулом была цифра «1» или «2». Участнику требовалось как можно быстрее ответить левым средним пальцем, если стимул был «1», или левым указательным пальцем, если стимул был «2». Во второй контрольной задаче (КЗ2) целевыми стимулами были названия цветов (либо «Желтый», либо «Синий»), представленные в их согласованном цвете, и участникам нужно было ответить правым указательным пальцем на «Желтый» или правым средним пальцем на «Синий». В КЗ3 участники снова видели названия цветов в согласованных цветах, но использовали «Красный», «Зеленый», «Желтый» и «Синий». Задача требовала от участников ответить левым средним пальцем на «Красный», левым указательным пальцем на «Зеленый», правым указательным пальцем на «Желтый» или правым средним пальцем на «Синий» (то есть во всех условиях «Желтый» и «Синий» всегда были сопоставлены с одними и теми же пальцами). Для всех трех контрольных задач в соответствующих блоках предъявлялось шесть испытаний.
Задачи исполнительных функций
Задача обновления (ОБН) была основана на задаче 1-назад. Стимулы и сопоставления клавиш были идентичны КЗ3, и участникам нужно было запомнить цвет, представленный в предыдущем испытании, и ответить на него. Для оценки специфической для ОБН активации мы рассчитали контраст ОБН – КЗ3 [1], поскольку в данном контексте КЗ3 представляла собой условие 0-назад.
Задача ингибирования (ИНГ) была основана на задаче Струпа. Стимулы и сопоставления клавиш были идентичны КЗ3, за исключением того, что названия цветов были представлены в несогласующемся цвете (например, «Красный» написано зеленым). Участникам было поручено как можно быстрее отвечать в зависимости от цвета каждого стимула. Другими словами, им нужно было игнорировать написанное слово и сосредоточиться на цвете шрифта. Для оценки специфической для ИНГ активации мы рассчитали контраст ИНГ – КЗ3 [1], поскольку в данном контексте КЗ3 представляла собой согласованное условие Струпа, эквивалентное условию чтения слов в традиционной бумажной задаче.
Выполнение двойной задачи (ДЗ). Испытания ДЗ представляли собой комбинацию цифр (как в КЗ1) и названий двух цветов (КЗ2). Одно испытание могло, например, одновременно предъявлять цифру «1» и название цвета «Синий», требуя двух ответов. Участникам нужно было отвечать сначала на цифру, а затем на название цвета соответственно (цифра → цвет). Поскольку две задачи относятся к одной и той же области (визуальные задачи), изменение порядка задач не привело бы к существенной разнице, поэтому мы всегда использовали один и тот же порядок задач. Оба стимула предъявлялись одновременно, то есть асинхрония стимулов (АС) всегда составляла 0 мс, поскольку мы хотели сравнить более высокие исполнительные требования (т.е. двойная задача с АС 0) с контрольными задачами, которые имеют незначительные исполнительные требования (одиночные задачи). Стимулы были взяты из всех комбинаций цифр и названий цветов: 1 Желтый, 2 Желтый, 1 Синий, 2 Синий. В каждом блоке предъявлялось десять испытаний в случайном порядке. В отличие от всех других задач, где в каждом испытании требовался один ответ, в задаче двойного выполнения требовалось два ответа за испытание. Для изучения специфической для выполнения двойной задачи активации, на первом уровне статистики, мы рассчитали контраст ДЗ – КЗ1 – КЗ2, то есть [1 -1 -1] индивидуально для каждого участника. Причина этого заключалась в том, что КЗ1 и КЗ2 составляли одиночные задачи, из которых состояла двойная задача. Следовательно, этот контраст выявлял бы специфические для двойной задачи активации, которые не могут быть объяснены суммированной активацией одиночных задач.
В задаче переключения (ПЕР) предъявлялись те же стимулы, что и для ДЗ (то есть «1 Желтый», «2 Желтый», «1 Синий», «2 Синий»), либо все черным цветом, либо в их согласованном цвете. Следовательно, в этой задаче было 8 стимулов (4 черных и 4 цветных варианта одних и тех же стимулов). Если стимулы были черными, участникам нужно было отвечать на основе отображаемых цифр (1 или 2) и отвечать так же, как описано для КЗ1. Если стимулы предъявлялись в цвете, участникам нужно было отвечать на основе цвета (Желтый или Синий), как описано для КЗ2. Поскольку слово и цвет были согласованы, участники могли основывать свое решение на семантике или цвете чернил (или обоих) слова. Подсказки «число» и «цвет» предъявлялись в случайном порядке, так что участники переключали свой ответ с одной размерности (число или цвет) на другую в зависимости от подсказки. Сопоставления ответов для цифр и цветов, а также параметры времени стимулов были идентичны параметрам выполнения двойной задачи. Для оценки специфической для ПЕР активации мы рассчитали контраст ПЕР – (КЗ1 + КЗ2)/2 [1 -0.5 -0.5]. Опять же, КЗ1 и КЗ2 составляли «компонентные» задачи ПЕР, но поскольку в ПЕР они выполняют только одну задачу за испытание, мы вычли среднее значение.
Процедура МРТ
Изображения мозга были получены с помощью 3 Т МРТ-сканера (Magnetom Trio, Siemens, Erlangen, Германия), оснащенного 32-канальной головной катушкой. 35 осевых срезов (поле зрения 192 × 192 мм, матрица 64 × 64, разрешение в плоскости 3 × 3 мм, толщина 3 мм, без зазора, чередующийся сбор срезов) были получены с использованием BOLD-чувствительной градиентно-эхо EPI последовательности (TR 2,5 с, TE 31 мс, угол наклона 85°). Высокоразрешающие изображения всего мозга были получены от каждого участника с использованием T1-взвешенной последовательности MPRAGE (TR 1900 мс, TE 3,03 мс, угол наклона 11°, 176 срезов, поле зрения 256 × 256 мм, размер вокселя 1 × 1 × 1 мм). Было получено функциональное сканирование с 755 томами, при этом выборка томов охватывала все 35 срезов.
Анализ данных
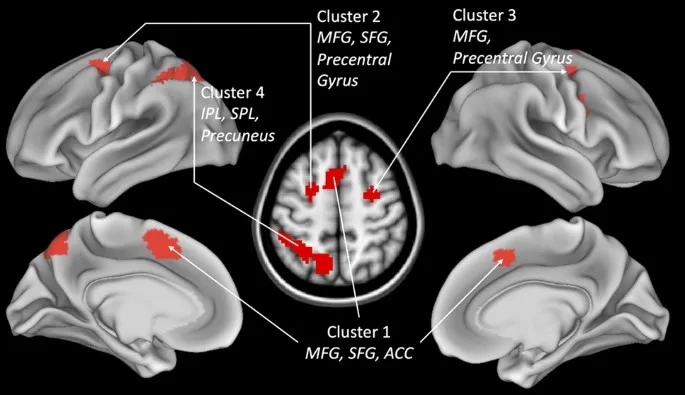
Мы использовали SPM12 (www.fil.ion.ucl.ac.uk/spm/) для анализа данных МРТ. Сначала мы вручную выровняли начало структурных и функциональных изображений с передней спайкой. Движение головы было скорректировано с помощью опции «Realign & Unwarp», и все изображения были трансформированы в пространство MNI с использованием опций нормализации и унифицированной сегментации. Наконец, мы сгладили изображения с помощью гауссова ядра с шириной на полувысоте (FWHM) 8 мм. Изображения были визуально проверены и валидированы относительно успешности нормализации и регистрации. Статистический анализ основывался на попиксельной оценке наименьших квадратов с использованием общей линейной модели для серийно автокоррелированных наблюдений. Поскольку в данном исследовании использовался блочный дизайн фМРТ, для моделирования BOLD-ответа использовалась функция boxcar, свертки с канонической HRF без производных. Для обеспечения возможности сравнения между задачами ИФ (длительностью 25 с) и контрольными задачами (длительностью 15 с) мы анализировали только первые 15 с каждого блока для всех условий (другими словами, при спецификации дизайна, начало всегда устанавливалось в начале блока, а параметр длительности всегда устанавливался на 15 с). Применялся временной высокочастотный фильтр с частотой среза 1/128 Гц. Были рассчитаны индивидуальные карты контрастов для всех контрастов интереса (см. раздел «Результаты»), а анализ второго уровня основывался на одновыборочных t-тестах. Для проверки областей разнообразия (т.е. областей, демонстрирующих различные паттерны активации для четырех ИФ) в функциональных нейроанатомических коррелятах четырех исполнительных функций мы рассчитали контрасты взаимодействия между всеми парами исполнительных функций. Эти контрасты взаимодействия сравнивали, различались ли области, специфичные для ИФ, путем вычитания соответствующих контрольных задач, а затем сравнения результатов этих вычитаний. Например, чтобы проверить, активируют ли обновление и ингибирование разные области, мы рассчитали [(ОБН – КЗ3) – (ИНГ – КЗ3)] и обратный [(ИНГ – КЗ3) – (ОБН – КЗ3)]. Для исследования общих активаций между четырьмя задачами ИФ мы провели конъюнктивный анализ с использованием логического подхода «И». Для конъюнктивного анализа мы использовали указанные выше контрасты (см. раздел 2.2.2, например, ИНГ – КЗ3), с порогом p < 0,194, предполагая, что нет значимых различий в силе активации между четырьмя ИФ в областях единства, показанных в таблице 1 (дополнительный рис. 2). Поэтому мы считаем эти четыре кластера областями истинного единства, которые не демонстрируют даже относительного разнообразия.
Разнообразие: различия между четырьмя ИФ
После определения областей единства мы далее проверили различия в функциональных нейроанатомических коррелятах четырех ИФ. Для этого мы рассчитали контрасты взаимодействия для парных сравнений четырех исполнительных функций (Таблица 2; Дополнительный рис. 3). Обратите внимание, что эти контрасты взаимодействия не могут отличить абсолютное и относительное разнообразие, и это различие будет сделано ниже.
Обновление
Сравнение обновления с выполнением двойной задачи [(ОБН – КЗ3) – (ДЗ – КЗ1 – КЗ2)] показало значительные активации в латеральной префронтальной, теменной, а также подкорковой областях. Кластер в латеральной префронтальной коре в основном охватывал двусторонние средние лобные извилины (BA 8), простираясь в правую верхнюю лобную извилину (BA 8). Кластер в теменной коре охватывал левую надкраевую извилину (BA39), простираясь в левую нижнюю теменную долю (BA39). Наконец, другой кластер в медиальных теменных областях располагался в основном в BA 31, охватывая правую заднюю поясную кору, простираясь в парацентральную долю и левую предклинье. При сравнении обновления с переключением [(ОБН – КЗ3) – (ПЕР – (КЗ1 + КЗ2)/2)] мы наблюдали три значительных кластера в лобных и теменных областях. Кластер в префронтальной коре в основном охватывал левую среднюю лобную извилину (BA 8/6), простираясь в правую среднюю лобную извилину (BA 6/8) и верхнюю лобную извилину (BA 6). Было два значительных кластера в теменных областях: один расположен в правой надкраевой извилине (BA 39/40), а другой — в правой задней поясной коре (BA 31), простираясь в левое предклинье (BA 7). Наконец, повышенная активация для обновления по сравнению с ингибированием [(ОБН – КЗ3) – (ИНГ – КЗ3)] выявила значительный кластер в правой латеральной префронтальной коре, простираясь от верхней лобной (BA 8) извилины к средней лобной извилине (BA 8). Таким образом, в основном префронтальные (СЛИ и ВЛИ) и теменные области (надкраевая извилина, простираясь к задней поясной коре) были более сильно связаны с обновлением, чем с другими тремя задачами исполнительных функций.
Ингибирование
Сравнение ингибирования с выполнением двойной задачи [(ИНГ – КЗ3) – (ДЗ – КЗ1 – КЗ2)] показало значительные активации в латеральных префронтальных областях, простираясь от левой средней лобной извилины (BA 6) к левой нижней лобной извилине (BA 46) и правой верхней лобной извилине (BA 9/8). Второй кластер располагался в двусторонних задних поясных коре (BA 31), простираясь к левому предклинью (BA 31). Сравнение ингибирования с переключением и обновлением не выявило значительных активаций (p > 0,005, не скорректировано).
Переключение
Сравнение переключения с ингибированием [(ПЕР – (КЗ1 + КЗ2)/2) – (ИНГ – КЗ3)] показало значительные активации в правой нижней (BA 40) и правой верхней теменной долях (BA 7). При сравнении с выполнением двойной задачи [(ПЕР – (КЗ1 + КЗ2)/2) – (ДЗ – КЗ1 – КЗ2)] результаты показали три значительных кластера в латеральной префронтальной, теменной и затылочной коре. Значительный кластер в латеральной префронтальной коре простирался от левой СЛИ (BA 6) к двусторонним верхним лобным извилинам (BA 8). Кластер в теменной области располагался вдоль двусторонних задних поясных извилин (BA 31). Наконец, в затылочной коре значительные активации располагались в двусторонних клиновидных долях (BA 18/19). Значительных кластеров при сравнении переключения с обновлением не обнаружено. Таким образом, в основном правополушарные корковые области, охватывающие верхнюю и нижнюю теменные доли, а также двусторонние полушарные корковые области, простирающиеся от левой СЛИ до двусторонних ВЛИ, простираясь к задней поясной коре и клиновидной доле, были более тесно связаны с процессами переключения, чем с ингибированием и выполнением двойной задачи.
Выполнение двойной задачи
Сравнение выполнения двойной задачи с переключением [(ДЗ – КЗ1 – КЗ2) – (ПЕР – (КЗ1 + КЗ2)/2)] выявило два значительных кластера с более сильной активацией для выполнения двойной задачи, чем для переключения: один во фронтотеменных областях и один в подкорковых областях. Большой кластер во фронтотеменной области в основном охватывал корковую область между правой постцентральной извилиной (BA 40) и правой средней лобной извилиной (BA 6), но сзади также простирался в левую постцентральную извилину (BA 3). Кластер в подкорковой области в основном охватывал таламус (BA 50) двусторонне. При сравнении выполнения двойной задачи с ингибированием [(ДЗ – КЗ1 – КЗ2) – (ИНГ – КЗ3)] мы наблюдали аналогичную закономерность результатов во фронтотеменной области, с кластером, простирающимся от правой постцентральной извилины (BA 40) до правой средней лобной извилины (BA 6). Результаты сравнения выполнения двойной задачи с обновлением [(ДЗ – КЗ1 – КЗ2) – (ОБН – КЗ3)] показали значительно увеличенные активации, простирающиеся от двусторонних постцентральных извилин (BA 40) до двусторонних нижних теменных долей (BA 40) и двусторонних верхних лобных извилин (BA 6). Таким образом, в основном правополушарные корковые области, простирающиеся от постцентральной извилины и нижней теменной доли к средним и верхним лобным извилинам, были более тесно связаны с процессами выполнения двойной задачи, чем с другими тремя исполнительными функциями.
Наконец, мы извлекли бета-значения в сфере радиусом 5 мм вокруг выбранных пиковых координат, идентифицированных в контрастах взаимодействия (Таблица 2), и проверили наличие активности в этих координатах для четырех контрастов, специфичных для ИФ [например, (ИНГ – КЗ3) для ингибирования]. Пики в непосредственной близости не рассматривались индивидуально, вместо этого был выбран наиболее значимый пик на рис. 3. Мы проверили одновыборочным t-тестом, значительно ли отличались средние бета-значения от нуля для каждой задачи ИФ. Если они отличались от нуля более чем для одной задачи, мы рассматривали это как область относительного разнообразия (рис. 3). Если они отличались только для одной задачи, мы рассматривали эту область как область абсолютного разнообразия (рис. 4). Области, не показавшие активации выше нуля, были исключены.
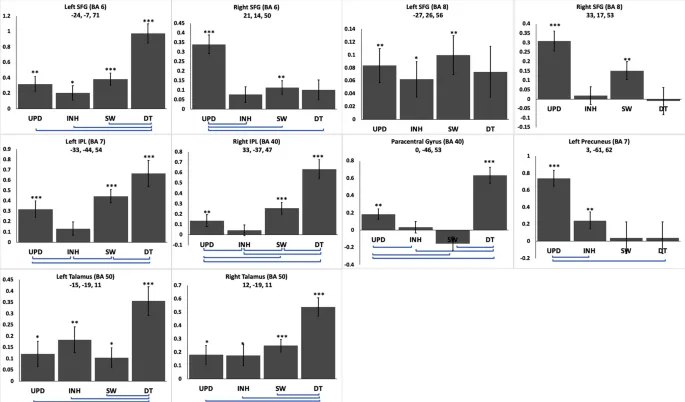
Результаты показали, что определенные области (т.е. левая ВЛИ (BA 6; координаты MNI x = -24, y = -7, z = 71) и двусторонний таламус (BA 50; -15, -19, 11 и 12, -19, 11) активировались для всех ИФ (все t (28) = t > 2,10, все p < 0,05, скорректировано).
Далее, были области, демонстрирующие значительную положительную активацию только для двух или трех ИФ. Левая НТД (BA 7) и правая НТД (BA 40) были активны только при ДЗ, ПЕР и ОБН. Левая ВЛИ (BA8) показала аналогичную значительную активацию для ОБН, ПЕР и ИНГ. Правая ВЛИ (BA6/8) (ОБН и ПЕР) и левое предклинье BA7 (ОБН и ИНГ) были значительно активированы только для двух ИФ. Правая парацентральная извилина показала активацию при ДЗ и ПЕР (рис. 4).
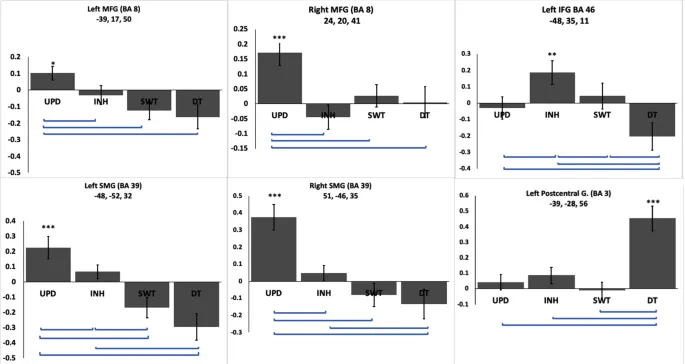
Наконец, наши результаты показали области абсолютного разнообразия, то есть области, которые были значительно активированы только в одной ИФ. В деталях, двусторонние СЛИ (BA 8) и двусторонние НКИ (BA39) были активны только во время обновления, левая НЛИ (BA 46) была активна только во время ингибирования, левая постцентральная извилина (BA 40) была активна только во время выполнения двойной задачи, в то время как областей абсолютного разнообразия для переключения не было (рис. 4).
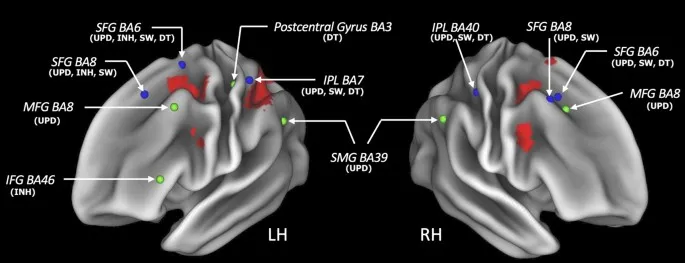
Взятые вместе, мы представили иллюстрацию областей, соответствующих относительному и абсолютному разнообразию, в дополнение к областям единства, на рис. 5.
Обсуждение
В данном исследовании мы стремились изучить единство и разнообразие нейронных коррелятов основных исполнительных функций: обновления, ингибирования, переключения и выполнения двойной задачи. Мы разработали четыре целевые задачи, которые были выведены из одной и той же базовой парадигмы, по одной для обновления, ингибирования, переключения и выполнения двойной задачи, а также три контрольные задачи. Наши поведенческие результаты показали, что участники были медленнее в каждой задаче ИФ по сравнению с соответствующей контрольной задачей. Аналогичные анализы в форме контрастов фМРТ (задачи ИФ – контрольные задачи) выявили активации в префронтальной и теменной коре для всех четырех ИФ. Затем мы охарактеризовали эти области как демонстрирующие единство (т.е. области, активированные всеми четырьмя ИФ), относительное разнообразие (т.е. области, активированные двумя или тремя ИФ) или абсолютное разнообразие (т.е. области, активированные только одной ИФ) (рис. 5).
Области единства располагались в основном в преимущественно леволатерализованных фронтотеменных областях, состоящих из медиальной префронтальной коры (МПФК), передней поясной коры (ППК), прецентральной извилины, верхней лобной извилины (ВЛИ), средней лобной извилины (СЛИ) (двусторонне), а также левой нижней и верхней теменных долей и предклинья. Области левого полушария могут быть более доминирующими в реализации ИФ, основываясь на результатах эмпирических исследований, а также на исследовании поражений. Например, два эмпирических исследования изучали ряд исполнительных функций, и оба исследования в целом показали, что левое полушарие связано с единством ИФ. В соответствии с этим, исследование поражений продемонстрировало, что пациенты с левым лобным повреждением показали худшие результаты, чем пациенты с правым лобным повреждением, в задаче «Башня Ханоя», которая связана с ИФ, такими как ингибирование и планирование. В своем основополагающем поведенческом исследовании Мияке и коллеги выделили общий фактор среди обновления, ингибирования и переключения, отражающий общий ментальный механизм ИФ, лежащий в основе различных задач ИФ. Мы интерпретируем области мозга, демонстрирующие единство, как связанные с этим общим механизмом ИФ.
Мы обнаружили лишь немногие области абсолютного разнообразия, то есть области, которые были связаны только с одной ИФ, но не с какой-либо другой, которые располагались в двусторонних СЛИ (BA 8) и левой надкраевой извилине (BA 39) для обновления, в левой НЛИ (BA 46) для ингибирования и в левой постцентральной извилине (BA 40) для выполнения двойной задачи. Больше областей показали относительное разнообразие, то есть связь с двумя или тремя ИФ, таких как двусторонние ВЛИ (BA 6, 8), правая СЛИ (BA 6), правая НТД (BA 40), левая ВТД (BA 7), парацентральная извилина (BA 40), двусторонний таламус (BA 50) и предклинье (BA 7). Эти результаты также отражают корреляционные закономерности между ИФ, выявленные Мияке и коллегами, которые обнаружили, что некоторая вариативность может быть объяснена парными корреляциями только двух ИФ, и что некоторая вариативность была уникальной для каждой ИФ. Следовательно, мы интерпретируем эти области абсолютного и относительного разнообразия как связанные с ментальными требованиями, специфичными для некоторых или даже только одной ИФ. Взятые вместе, используя внутрисубъектный дизайн, мы обнаружили, что четыре исследованные ИФ частично обслуживаются общим нейронным основанием (единство) и частично — различными, специфичными для ИФ, нейронными коррелятами (разнообразие).
Мы обнаружили, что области единства были организованы во фронтотеменных областях, что согласуется с исследованием Коллетт и коллег, которые также сообщали об областях единства в теменных и лобных областях, хотя лобные области были очевидны только при сниженном пороге, и с Деррфусом и коллегами, которые нашли аналогичную закономерность результатов в настоящем исследовании. В целом, ряд исследований показал, что ИФ связаны с фронтотеменной сетью, и что лобные области (такие как ВЛИ, СЛИ и МПФК) функционально связаны с теменными областями (такими как НТД и ВТД), обеспечивая высшие психические операции, такие как реализация целенаправленного поведения, выбор ответов, поддержание памяти и разрешение конфликтов. Ни и Браун продемонстрировали, что задняя часть левой лобной коры (BA 6) связана с левыми задними теменными областями, и что эти лобные и теменные доли активируются широким спектром задач ИФ. Эта идея о фронтотеменных областях, обслуживающих высшие когнитивные операции, согласуется с точкой зрения на контроль внимания, предложенной Энглом и коллегами, в которой ИФ относятся к общей и доменно-независимой способности, не связанной исключительно с конкретными задачами. Взятые вместе, представляется вероятным, что эти фронтотеменные области, связанные с единством ИФ, могут действовать как центральный узел контроля внимания, интегрируя входные данные, такие как представление правил задачи из различных корковых областей, и осуществляя свой контроль путем манипулирования нейронной обработкой в областях, связанных с более специфической обработкой, связанной с задачей.
Кроме того, предыдущие исследования предложили две возможные причины перекрытия во всех трех ИФ. Во-первых, «теория разделения» предположила, что определенные области мозга могут быть вовлечены в обработку всех задач. Эта идея «теории разделения» была интерпретирована двумя способами. «Редукционистская интерпретация» утверждает, что общая область может быть связана с одной задачей и может быть заимствована другой задачей. То есть, относительно простая задача активирует область, которая также задействована в более сложной задаче, активация в сложной задаче может отражать основное требование простой задачи и быть заимствована последней задачей. «Абстрактивная интерпретация» предполагает, что общие активности в мозге должны описываться более абстрактными терминами, а не приписываться одной задаче. Например, общие области могут быть связаны с более общими операциями, такими как мониторинг, которые могут быть задействованы во всех ИФ. Во-вторых, «сетевая теория» утверждает, что определенные области мозга могут активироваться для всех задач, но это не обязательно означает, что они связаны с одной и той же когнитивной операцией, потому что общие области могут быть функционально связаны с другими областями мозга, которые могут помочь выполнять различные операции. Было указано, что эти теории не противоречат друг другу, а скорее могут комбинироваться различными способами. В этом отношении представляется вероятным, что абстрактивная интерпретация более согласуется с нашими выводами относительно единства четырех ИФ. Причина этого в том, что для минимизации влияния задач мы создали четыре различные версии, основанные на одной и той же базовой задаче, которые основаны на одних и тех же стимулах и в основном отличаются инструкциями, данными участникам. Поскольку редукционистская интерпретация обычно используется для перекрытия между двумя или тремя задачами, мы применяем ее для интерпретации относительного разнообразия ниже.
В текущем исследовании мы обнаружили единство нейронной основы ИФ не только для переключения, ингибирования и обновления, но и для выполнения двойной задачи. Мияке и коллеги предположили, что три задачи ИФ (т.е. переключение, ингибирование и обновление) коррелировали друг с другом с точки зрения поведенческой производительности, но не наблюдали корреляций с выполнением двойной задачи. В то время как Мияке и коллеги предположили, что процесс двойной задачи может требовать независимой исполнительной функции, наши нейровизуализационные данные предполагают, что выполнение двойной задачи разделяет нейронные корреляты с другими тремя ИФ. Потенциальной причиной этого расхождения может быть использование различных парадигм двойной задачи. Мияке и коллеги использовали довольно сложную парадигму двойной задачи, состоящую из двух различных задач рабочей памяти (тест скорости прохождения лабиринта и генерация букв), для которых точная временная структура ментальных процессов трудно установить. В нашей задаче мы объединили две задачи быстрого выбора ответа, предъявляемые одновременно, для которых хорошо известно, что обе задачи конкурируют за ограниченный ресурс обработки. Наши выводы о том, что выполнение двойной задачи также активирует области единства, согласуются с поведенческими исследованиями, теоретическими моделями, а также нейровизуализационными исследованиями, предполагающими, что выполнение двойной задачи требует ИФ для координации одновременной обработки двух задач. Таким образом, наше исследование предполагает наличие единства в нейронных коррелятах четырех различных ИФ.
Области относительного разнообразия показали, что ИФ совместно активировали определенные области в обоих полушариях с различной силой. В соответствии с редукционистской точкой зрения, это наблюдение может указывать на то, что определенные ИФ могут частично перекрываться в разной степени по своим ментальным требованиям через общие требования задачи. Например, вполне вероятно, что большинство задач переключения также требуют ингибирования, поскольку для успешной операции переключения задача, от которой нужно переключиться, может потребовать подавления. Аналогично, ряд исследований согласились с тем, что обработка двойной задачи включает ингибирование, переключение и обновление.
Мы обнаружили области абсолютного разнообразия, то есть области, которые были значительно активированы только одной ИФ, для ингибирования, обновления и выполнения двойной задачи. Эти области согласуются с предыдущими исследованиями, связывающими их с соответствующими ИФ. Например, НЛИ неоднократно признавалась ключевой областью ингибирования, а выполнение двойной задачи показало нарушение при повреждении левой постцентральной извилины (BA 40). Наконец, обновление было связано с двусторонними СЛИ (BA 8) и левой НКИ (BA 39) потенциально из-за требований к активному мониторингу. Мы не обнаружили области абсолютного разнообразия для переключения. Является ли это следствием недостаточной статистической мощности, особенностей задачи или того, что переключение не связано ни с какими уникальными областями, в рамках данного исследования ответить невозможно.
Мы хотели бы отметить, что наши обсуждения относительного и абсолютного разнообразия относятся к четырем ИФ, исследованным в данном исследовании. Мы не исключаем, что эти области активируются другими ИФ, не исследованными здесь, или даже другими ментальными процессами, не связанными с исполнительными функциями. Другими словами, мы не считаем их исключительно связанными с исследованными здесь функциями. Следовательно, область, которую мы считали демонстрирующей абсолютное разнообразие, потому что она была активна только в одной ИФ в этом исследовании, может быть активирована другой ИФ, например, планированием, и тогда может рассматриваться как область относительного разнообразия.
В анализе относительного разнообразия, в котором мы анализировали бета-значения всех четырех ИФ, мы определили области, которые активировались всеми четырьмя ИФ, хотя они не были идентифицированы в конъюнктивном анализе, конкретно тестирующем единство (например, таламус (BA 50) для выполнения двойной задачи). Однако анализ бета-значений был более чувствительным (нескорректированный p < 0,05) по сравнению с конъюнктивным анализом (скорректированный уровень кластера p < 0,05). Обоснованием использования различных порогов было то, что при анализе бета-значений мы хотели убедиться, что мы не ошибочно примем область за область абсолютного разнообразия, хотя она может демонстрировать некоторую активность в других областях (т.е. мы стремились исключить бета-ошибки). С другой стороны, в конъюнктивном анализе мы хотели убедиться, что мы идентифицируем только те области, которые действительно были активированы четырьмя ИФ (т.е. мы стремились исключить альфа-ошибки). Следовательно, использование различных порогов может объяснить такие наблюдения. В целом, точный пространственный охват активаций зависит от выбранных порогов. Таким образом, особенно на краях кластеров активации, где области единства могут плавно переходить в области относительного разнообразия, точное расположение этих переходов, вероятно, будет зависеть от статистической мощности и статистических порогов. Поэтому следует проявлять осторожность при интерпретации точного охвата областей единства, а также областей относительного и абсолютного разнообразия.
Настоящее исследование имеет некоторые присущие ограничения, которые следует учитывать в будущих исследованиях. Во-первых, длина блока для задач ИФ (25 с) и контрольных задач (15 с) была разной из-за непреднамеренной ошибки во время разработки эксперимента. Представленный здесь анализ был взят из первых 15 с каждого блока (см. «Методы»), но мы также провели идентичный анализ, используя полную длину блока, и это выявило очень похожую закономерность результатов. Во-вторых, задачи внутри каждого блока предъявлялись в повторяющемся (но рандомизированном) порядке. Хорошо известно, что повторение задач может привести к эффектам привыкания, поскольку участники привыкают к задаче. Мы изучили время ответа на задачи, и в целом, казалось, не было значительных изменений во времени ответа между испытаниями. На этом основании мы полагаем, что эффекты привыкания в наших данных маловероятны, но будущие исследования должны учитывать эту возможность. В-третьих, наша двойная задача предусматривала, что участники должны были отвечать в определенном порядке, а не иметь возможность отвечать в любом предпочитаемом ими порядке. Это может вызвать активацию областей мозга, отличных от тех, которые связаны с выполнением двойной задачи без какого-либо требуемого порядка, например, путем введения дополнительных требований к рабочей памяти. Наконец, анализ наших данных нейровизуализации использовал стандартный размер сглаживающего ядра, который неизбежно имеет последствия для идентификации и интерпретации паттернов активации (по мере увеличения ширины фильтра пространственный охват активации увеличивается). Чувствительны ли идентифицированные здесь области, связанные с единством и разнообразием, к размеру ядра, неясно, но, учитывая относительную близость некоторых из этих областей, это нельзя исключить.
В заключение, данное исследование было направлено на изучение нейронных коррелятов четырех исполнительных функций: переключения, ингибирования, обновления и выполнения двойной задачи. Мы обнаружили, что определенные области во фронтотеменной коре в целом активировались всеми четырьмя ИФ, и предполагаем, что эти области связаны с единой основной функцией, такой как управляемое внимание. Мы интерпретируем области, которые были активированы одной, двумя или тремя ИФ, как связанные с более специфическими ментальными требованиями, связанными только с некоторыми ИФ. Наши данные показывают, что ранее предложенная организация ИФ на единство и разнообразие также отражена в их функциональных нейроанатомических коррелятах.